B. 細菌のDNA複製
一般にDNAの複製は、らせんの巻き戻し、相補鎖間の水素結合の切断による鎖の分離、相補的塩基対形成による新しい2本の鎖の合成によって行われます。
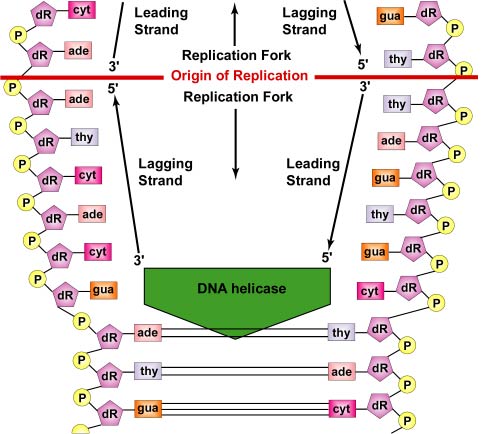
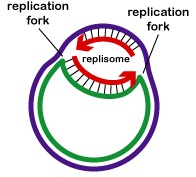
DNA 複製は、複製起点から双方向に行われます。 DNAの複製を開始するには、DNAヘリカーゼと呼ばれる巻き戻し酵素が、2本の親DNA鎖の短いセグメントを巻き戻し、複製起点で互いに分離させて、2つの「Y」字型の複製フォークを形成させる。 この複製フォークは、実際にDNAがコピーされる場所である(図参照)。
一本鎖結合タンパク質は一本鎖領域に結合し、二本鎖が再び結合しないようにします。 二本鎖らせんの巻き戻しは、複製分枝の前方に正のスーパーコイルを発生させる。 トポイソメラーゼと呼ばれる酵素は、DNAに切断を生じさせ、それを再結合して負のスーパーコイルを形成することでこれに対抗し、複製中のらせん分子におけるこのストレスを和らげる。
鎖がほぐれ続け、DNA分子全体の周りで両方向に分離すると、自由なDNAヌクレオチドがそれぞれの親鎖上のヌクレオチドと水素結合することによって、新しい相補鎖が作られる。 新しいヌクレオチドが水素結合によってそれぞれの親鎖の反対側に並ぶと、DNAポリメラーゼと呼ばれる酵素がホスホジエステル結合によってヌクレオチドを結合させる。 実は、相補的な塩基対によって並ぶヌクレオチドは、窒素塩基とデオキシリボースと3つのリン酸塩からなるデオキシヌクレオチド3リン酸塩である。 新しいヌクレオチドの5’リン酸基とDNA鎖の最後のヌクレオチドの3’OH基の間でホスホジエステル結合が形成されると、リン酸塩のうち2つが取り除かれて結合のためのエネルギーとなります(図参照)。 そして、それぞれの親鎖を鋳型として、相補的な自分自身のコピーを合成し、2つの同じDNA分子が形成されます(図(㊦))。 細菌では、Parタンパク質が、細胞分裂の際に細菌の染色体を細胞の反対側の極に分離する働きをします。 真核細胞の分裂装置と同様に、DNAの複製起点に結合して、染色体を物理的に引き離したり、押したりする。
ディビソームのFtsKなどのFtsタンパク質も、複製された細菌染色体を分離するのに役立ちます。
相補的塩基対によるDNA複製のGIFアニメーション
実際には、DNA複製はDNAポリメラーゼの性質によりこれより複雑です。 DNAポリメラーゼ酵素は、新しいヌクレオチドの5’炭素のリン酸基を、すでに鎖にあるヌクレオチドの3’炭素のヒドロキシル(OH)基と結合させることしかできないのである。 その結果、DNAは3’から5’方向に走る親鎖をコピーしながら、5’から3’方向にのみ合成することができる。
各DNA鎖には2つの末端があります。 5’末端はデオキシリボースの5’炭素に末端のリン酸基がある方で、3’末端はデオキシリボースの3’炭素に末端の水酸基(OH)がある方です(図⑷参照)。 この2本の鎖は反平行、つまり反対方向に走っている。 したがって、一方の親鎖(3’から5’を走るもの、リーディングストランドと呼ばれる)は、その全長をそのままコピーすることができる(図参照)。 しかし、もう一方の親鎖(5’から3’を走る、遅行鎖と呼ばれる)は、DNAが巻き戻される際に、約100-1000ヌクレオチドの短い断片(岡崎フラグメント)に不連続にコピーされなければならない。 これは、前述したように、レプリソームで起こる。 遅れているDNA鎖が先行する鎖からループ状になることで、レプリソームが両方の鎖に沿って移動し、複製が行われる際にDNAを引っ張り出すことができるのです。
さらに、DNAポリメラーゼ酵素は、ゼロから新しいDNA鎖を作ることはできません。
また、DNAポリメラーゼの酵素は、ゼロから新しいDNA鎖を作ることはできないが、既存の鎖にあるヌクレオチドの3’OH基の上に新しいヌクレオチドをつけることができるのみである。 したがって、先行鎖と遅発鎖の各DNA断片の合成を開始するためには、プライマーゼと呼ばれるRNAポリメラーゼ複合体が必要である。 既存の核酸鎖を必要とせず、RNAヌクレオチドを結合することができるプライマーゼは、まず親鎖のDNAヌクレオチドの反対側に、いくつかの相補的なRNAヌクレオチドを付加する。
次にDNAポリメラーゼIIIがプライマーゼに代わり、RNAプライマーにDNAを付加することができるようになります(図(ⅳ))。 その後、DNAポリメラーゼIIがRNAプライマーを消化し、プライマーのRNAヌクレオチドを適切なDNAヌクレオチドに置き換えて隙間を埋める(図(㊦))。 最後に、DNAリガーゼという酵素でDNA断片をつなぎ合わせる(図参照)。
DNA複製のアニメーション
HHMIのバイオインタラクティブによる提供
詳細はこちら(英語)。 原核生物のDNA複製の復習(ユニット7より)