- Objectivos de Aprendizagem
- B Produção e Maturação Celular
- Pense nisto
- B-Cell Receptors
- Pense nisso
- Activação das células B independente de T
- Pense nisso
- Ativação dependente da célula T de células B
- Pense em It
- Respostas primárias e secundárias
- Pense nisso
- Conceitos e Resumo Chave
- Selecção múltipla
- Fill in the Blank
- Think about It
Objectivos de Aprendizagem
- Descrever a produção e maturação das células B
- Comparar a estrutura dos receptores de células B e receptores de células T
- Comparar T-ativação dependente e independente em T das células B
- Comparar as respostas de anticorpos primários e secundários
Imunidadeumoral refere-se a mecanismos das defesas imunológicas adaptativas que são mediadas por anticorpos secretados por linfócitos B, ou células B. Esta secção irá focar as células B e discutir a sua produção e maturação, receptores e mecanismos de activação.
B Produção e Maturação Celular
Células T, as células B são formadas a partir de células estaminais hematopoiéticas multipotentes (HSCs) na medula óssea e seguem um caminho através de células estaminais linfóides e linfoblasto (ver Figura 1 nas Defesas Celulares). Ao contrário das células T, porém, os linfoblastos destinados a tornarem-se células B não deixam a medula óssea e viajam para o timo para amadurecer. Ao contrário, eventuais células B continuam a amadurecer na medula óssea.
O primeiro passo da maturação das células B é uma avaliação da funcionalidade dos seus receptores de ligação antigénica. Isto ocorre através da selecção positiva para as células B com receptores funcionais normais. Um mecanismo de selecção negativa é então utilizado para eliminar as células B auto-reactivas e minimizar o risco de auto-imunidade. A selecção negativa das células B auto-reactivas pode envolver a eliminação por apoptose, edição ou modificação dos receptores de modo a que deixem de ser auto-reactivas, ou indução de anergia na célula B. As células B imaturas que passam a selecção na medula óssea viajam então para o baço para as suas fases finais de maturação. Lá elas se tornam células B maduras ingênuas, ou seja células B maduras que ainda não foram activadas.
Pense nisto
- Compare a maturação das células B com a maturação das células T.
B-Cell Receptors
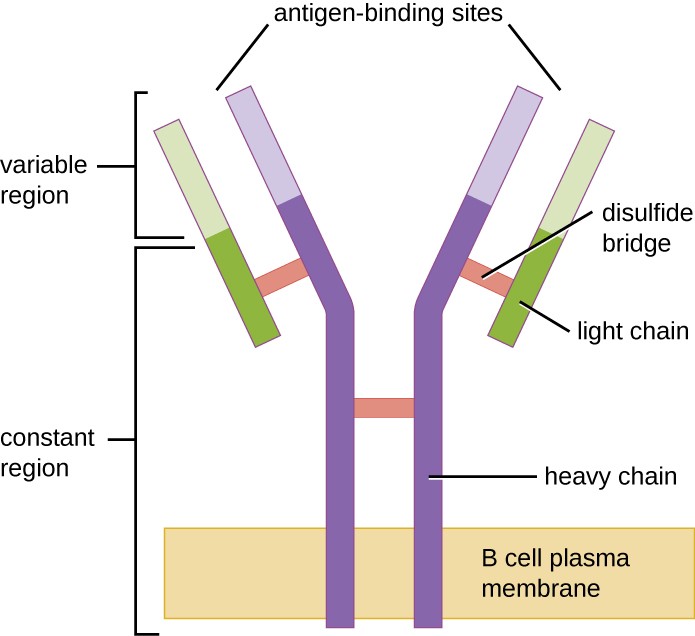
Figure 1. Os receptores das células B estão embutidos nas membranas das células B. As regiões variáveis de todos os receptores de uma única célula ligam o mesmo antígeno específico.
Células T, as células B possuem receptores específicos de antígenos com diversas especificidades. Apesar de dependerem das células T para uma função óptima, as células B podem ser activadas sem a ajuda das células T. Os receptores de células B (BCRs) para células B maduras ingênuas são formas monoméricas de IgD e IgM ligadas à membrana. Têm duas cadeias pesadas idênticas e duas cadeias leves idênticas ligadas por ligações de dissulfureto numa forma básica em “Y” (Figura 1). O tronco da molécula em forma de Y, a região constante das duas cadeias pesadas, atravessa a membrana da célula B. Os dois locais de ligação de antígenos expostos ao exterior da célula B estão envolvidos na ligação de epitopos patogénicos específicos para iniciar o processo de activação. Estima-se que cada célula B madura naïve tem mais de 100.000 BCRs na sua membrana, e cada um destes BCRs tem uma especificidade de ligação de epitopos idêntica.
Para estarem preparadas para reagir a uma vasta gama de epitopos microbianos, as células B, tal como as células T, usam o rearranjo genético de centenas de segmentos de genes para fornecer a diversidade necessária de especificidades de receptores. A região variável da cadeia pesada BCR é composta por segmentos V, D e J, semelhante à cadeia β do TCR. A região variável da cadeia leve do BCR é composta de segmentos V e J, semelhante à cadeia α do TCR. O rearranjo genético de todas as combinações possíveis de V-J-D (cadeia pesada) e V-J (cadeia leve) proporciona milhões de sítios únicos de ligação antigênica para o BCR e para os anticorpos segregados após a ativação.
Uma diferença importante entre BCRs e TCRs é a maneira como eles podem interagir com epitopos antigênicos. Enquanto os TCRs só podem interagir com epitopes antigênicos que são apresentados dentro da fenda de ligação de antígenos do MHC I ou MHC II, os BCRs não requerem apresentação de antígenos com MHC; eles podem interagir com epitopes em antígenos livres ou com epitopes apresentados na superfície de patógenos intactos. Outra diferença importante é que os TCRs só reconhecem epitopes de proteínas, enquanto os BCRs podem reconhecer epitopes associados a diferentes classes moleculares (por exemplo, proteínas, polissacarídeos, lipopolissacarídeos).
Ativação das células B ocorre através de diferentes mecanismos, dependendo da classe molecular do antígeno. A ativação de uma célula B por um antígeno proteico requer que a célula B funcione como um APC, apresentando os epitopos proteicos com MHC II para auxiliar as células T. Devido à sua dependência das células T para ativação das células B, os antígenos protéicos são classificados como antígenos dependentes de T. Em contraste, polissacarídeos, lipopolissacarídeos e outros antígenos não-proteicos são considerados antígenos T-independentes porque podem ativar células B sem processamento e apresentação de antígenos para células T.
Pense nisso
- Que tipos de moléculas servem como BCR?
- Quais são as diferenças entre TCRs e BCRs no que diz respeito ao reconhecimento de antígenos?
- Que classes de moléculas são antigénios dependentes de T e quais são antigénios independentes de T?
Activação das células B independente de T
Activação das células B sem a cooperação das células T de ajuda é referida como activação independente das células T e ocorre quando os BCRs interagem com antigénios independentes de T. Os antígenos independentes de T (por exemplo, cápsulas de polissacarídeo, lipopolissacarídeo) têm unidades epitopárias repetitivas dentro da sua estrutura, e esta repetição permite a ligação cruzada de múltiplos BCR, fornecendo o primeiro sinal para activação (Figura 2). Como as células T não estão envolvidas, o segundo sinal tem que vir de outras fontes, como interações de receptores tipo toll-like com PAMPs ou interações com fatores do sistema complemento.
Após uma célula B ser ativada, ela sofre proliferação clonal e as células filhas se diferenciam em plasmócitos. Células plasmáticas são fábricas de anticorpos que secretam grandes quantidades de anticorpos. Após a diferenciação, os BCRs de superfície desaparecem e o plasmócito secreta moléculas IgM pentamericanas que têm a mesma especificidade antigênica que os BCRs (Figura 2).
A resposta independente da célula T é de curta duração e não resulta na produção de células B de memória. Assim, não resultará numa resposta secundária a exposições subsequentes a antigénios independentes de T.

Figure 2. Os antigénios independentes do T têm epitopos de repetição que podem induzir o reconhecimento e activação das células B sem o envolvimento das células T. Um segundo sinal, como a interação de TLRs com PAMPs (não mostrado), também é necessário para a ativação da célula B. Uma vez ativada, a célula B prolifera e se diferencia em plasmócitos secretores de anticorpos.
Pense nisso
- Quais são os dois sinais necessários para a ativação das células T independente da célula B?
- Qual é a função de um plasmócito?
Ativação dependente da célula T de células B

Figure 3. Clique para uma imagem maior. Na ativação de células T dependente de células B, a célula B reconhece e internaliza um antígeno e o apresenta a uma célula T auxiliar que é específica para o mesmo antígeno. A célula auxiliar T interage com o antígeno apresentado pela célula B, que activa a célula T e estimula a libertação de citocinas que, por sua vez, activam a célula B. A ativação da célula B desencadeia a proliferação e diferenciação em células B e células plasmáticas.
A ativação dependente da célula T das células B é mais complexa que a ativação independente da célula T, mas a resposta imune resultante é mais forte e desenvolve a memória. A ativação dependente da célula T pode ocorrer tanto em resposta a antígenos protéicos livres quanto a antígenos protéicos associados a um patógeno intacto. A interação entre os BCRs em uma célula B naïve madura e um antígeno protéico livre estimula a internalização do antígeno, enquanto a interação com antígenos associados a um patógeno intacto inicia a extração do antígeno do patógeno antes da internalização. Uma vez internalizado dentro da célula B, o antígeno protéico é processado e apresentado com MHC II. O antígeno apresentado é então reconhecido pelas células T do helper específicas para o mesmo antígeno. O TCR da célula T do helper reconhece o antígeno estranho e a molécula CD4 da célula T interage com o MHC II na célula B. A coordenação entre as células B e as células T do ajudante que são específicas do mesmo antigénio é referida como reconhecimento ligado.
p>Após activado pelo reconhecimento ligado, as células TH2 produzem e secretam citocinas que activam a célula B e provocam a proliferação em células filhas clonais. Após vários ciclos de proliferação, citocinas adicionais fornecidas pelas células TH2 estimulam a diferenciação dos clones de células B ativadas em células B de memória, que responderão rapidamente às exposições subsequentes ao mesmo epitópo proteico, e plasmócitos que perdem seus BCRs de membrana e secretam inicialmente IgM pentamericano (Figura 3).
Após a secreção inicial de IgM, as citocinas secretadas por células TH2 estimulam os plasmócitos a passar da produção de IgM para a produção de IgG, IgA, ou IgE. Este processo, chamado mudança de classe ou mudança de isótipo, permite aos plasmócitos clonados a partir da mesma célula B activada produzir uma variedade de classes de anticorpos com a mesma especificidade de epitópo. A comutação de classe é realizada através do rearranjo genético de segmentos de genes que codificam a região constante, o que determina a classe de um anticorpo. A região variável não é alterada, portanto a nova classe de anticorpos mantém a especificidade do epitopo original.
Pense em It
- Que passos são necessários para a ativação das células B dependentes da célula T?
- O que é mudança de classe de anticorpos e por que é importante?
Respostas primárias e secundárias
Ativação dependente de célula T de células B desempenha um papel importante tanto nas respostas primárias quanto nas secundárias associadas à imunidade adaptativa. Com a primeira exposição a um antígeno proteico, ocorre uma resposta de anticorpos primários dependentes de células T. O estágio inicial da resposta primária é um período de atraso, ou período latente, de aproximadamente 10 dias, durante o qual nenhum anticorpo pode ser detectado no soro. Este período de retardamento é o tempo necessário para todas as etapas da resposta primária, incluindo a ligação ingenuamente madura das células B do antígeno com BCR, processamento e apresentação do antígeno, ativação da célula T auxiliar, ativação da célula B e proliferação clonal. O fim do período de atraso é caracterizado por um aumento dos níveis de IgM no soro, uma vez que as células TH2 estimulam a diferenciação das células B em plasmócitos. Os níveis de IgM atingem seu pico cerca de 14 dias após a exposição primária ao antígeno; mais ou menos ao mesmo tempo, o TH2 estimula a mudança de classe de anticorpos, e os níveis de IgM no soro começam a diminuir. Entretanto, os níveis de IgG aumentam até atingirem um pico de cerca de três semanas após a resposta primária (Figura 4).
Durante a resposta primária, algumas das células B clonadas são diferenciadas em células B de memória programadas para responder a exposições subsequentes. Esta resposta secundária ocorre mais rapidamente e com mais força do que a resposta primária. O período de atraso é reduzido para apenas alguns dias e a produção de IgG é significativamente maior do que a observada para a resposta primária (Figura 4). Além disso, os anticorpos produzidos durante a resposta secundária são mais eficazes e ligam-se com maior afinidade aos epitopos visados. As células plasmáticas produzidas durante as respostas secundárias vivem mais tempo do que as produzidas durante a resposta primária, portanto os níveis de anticorpos específicos permanecem elevados por um longo período de tempo.

Figure 4. Em comparação com a resposta primária, a resposta de anticorpos secundários ocorre mais rapidamente e produz níveis de anticorpos mais altos e mais sustentados. A resposta secundária envolve principalmente IgG.
Pense nisso
- Que eventos ocorrem durante o período de atraso da resposta primária de anticorpos?
- Por que os níveis de anticorpos permanecem elevados por mais tempo durante a resposta de anticorpos secundários?
Conceitos e Resumo Chave
- linfócitos B ou células B produzem anticorpos envolvidos na imunidade humoral. As células B são produzidas na medula óssea, onde ocorrem os estágios iniciais de maturação, e viajam até o baço para os estágios finais de maturação em células B maduras ingênuas.
- receptores de células B (BCRs) são formas monoméricas de IgD e IgM que ligam epitopos antigênicos específicos com suas regiões de ligação ao antígeno Fab. A diversidade da especificidade de ligação de antígenos é criada pelo rearranjo genético dos segmentos V, D e J, semelhante ao mecanismo usado para a diversidade de TCR.
- Antígenos proteicos são chamados antígenos dependentes de T porque só podem ativar células B com a cooperação de células T helper. Outras classes de moléculas não requerem cooperação de células T e são chamadas de antígenos T-independentes.
- activação de células B dependentes de células T envolve processamento e apresentação de antigénios proteicos para ajudar as células T, activação das células B por citocinas secretadas de células TH2 activadas e plasmócitos que produzem diferentes classes de anticorpos como resultado da mudança de classe. Células B de memória também são produzidas.
- As exposições secundárias a antígenos dependentes de T resultam em uma resposta secundária de anticorpos iniciada pelas células B de memória. A resposta secundária desenvolve-se mais rapidamente e produz níveis mais elevados e sustentados de anticorpos com maior afinidade para o antígeno específico.
li> A ativação de células B independente de células T envolve a ligação cruzada de BCRs por epitopes repetitivos de antígenos não-proteicos. Caracteriza-se pela produção de IgM por plasmócitos e não produz células B de memória.
Selecção múltipla
Qual dos seguintes seria um antígeno dependente de T?
- lipopolysaccharide
- glycolipid
- protein
- carbohydrate
Which of the following would be a BCR?
- CD4
- MHC II
- MHC I
- IgD
Which of the following does not occur during the lag period of the primary antibody response?
- activation of helper T cells
- class switching to IgG
- presentation of antigen with MHC II
- binding of antigen to BCRs
Fill in the Blank
________ antigens can stimulate B cells to become activated but require cytokine assistance delivered by helper T cells.
T-independent antigens can stimulate B cells to become activated and secrete antibodies without assistance from helper T cells. These antigens possess ________ antigenic epitopes that cross-link BCRs.
Think about It
A patient lacks the ability to make functioning T cells because of a genetic disorder. Would this patient’s B cells be able to produce antibodies in response to an infection? Explain your answer.