- Cele nauczania
- Produkcja i dojrzewanie komórek B
- Pomyśl o tym
- Receptory komórek B
- Pomyśl o tym
- Niezależna od komórek T aktywacja komórek B
- Pomyśl o tym
- Zależna od komórek T aktywacja komórek B
- Pomyśl o tym
- Pierwotna i wtórna odpowiedź
- Pomyśl o tym
- Kluczowe pojęcia i podsumowanie
- Wielokrotny wybór
- Fill in the Blank
- Think about It
Cele nauczania
- Opisać wytwarzanie i dojrzewanie komórek B
- Porównać strukturę receptorów komórek B i receptorów komórek T
- Porównać aktywację T-i T-niezależną aktywację komórek B
- Porównaj pierwotną i wtórną odpowiedź przeciwciał
Odporność humoralna odnosi się do mechanizmów adaptacyjnej obrony immunologicznej, w której pośredniczą przeciwciała wydzielane przez limfocyty B, lub komórki B. W tej sekcji skupimy się na komórkach B i omówimy ich produkcję i dojrzewanie, receptory i mechanizmy aktywacji.
Produkcja i dojrzewanie komórek B
Podobnie jak limfocyty T, komórki B powstają z multipotencjalnych krwiotwórczych komórek macierzystych (HSCs) w szpiku kostnym i podążają ścieżką poprzez limfoidalną komórkę macierzystą i limfoblast (patrz Rycina 1 w Obronie komórkowej). W przeciwieństwie do komórek T, limfoblasty, które mają stać się komórkami B, nie opuszczają szpiku kostnego i nie wędrują do grasicy w celu dojrzewania. Pierwszym etapem dojrzewania komórek B jest ocena funkcjonalności ich receptorów wiążących antygen. Następuje to poprzez selekcję pozytywną dla komórek B z normalnymi funkcjonalnymi receptorami. Następnie stosuje się mechanizm selekcji negatywnej w celu wyeliminowania samoreagujących komórek B i zminimalizowania ryzyka wystąpienia autoimmunizacji. Negatywna selekcja samoreagujących komórek B może obejmować eliminację poprzez apoptozę, edycję lub modyfikację receptorów, tak aby nie były już samoreaktywne, lub indukcję anergii w komórce B. Niedojrzałe komórki B, które przejdą selekcję w szpiku kostnym, wędrują następnie do śledziony, gdzie przechodzą końcowe etapy dojrzewania. Tam stają się naiwnymi dojrzałymi komórkami B, tzn, dojrzałymi komórkami B, które nie zostały jeszcze aktywowane.
Pomyśl o tym
- Porównaj dojrzewanie komórek B z dojrzewaniem limfocytów T.
Receptory komórek B
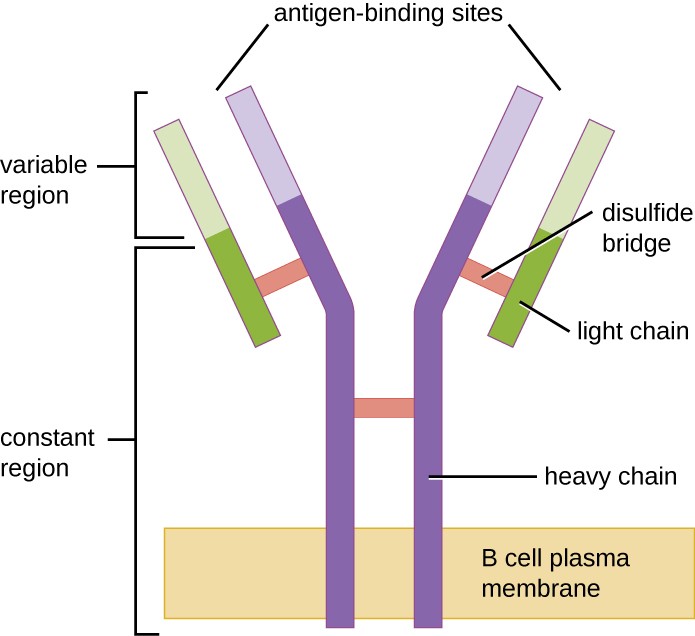
Rysunek 1. Receptory B-komórkowe są wbudowane w błony komórek B. Regiony zmienne wszystkich receptorów na jednej komórce wiążą ten sam specyficzny antygen.
Podobnie jak limfocyty T, limfocyty B posiadają receptory specyficzne dla antygenów o różnej specyficzności. Chociaż zależą one od komórek T dla optymalnego funkcjonowania, komórki B mogą być aktywowane bez pomocy komórek T. Receptory komórek B (BCR) dla naiwnych dojrzałych komórek B to związane z błoną monomeryczne formy IgD i IgM. Mają one dwa identyczne łańcuchy ciężkie i dwa identyczne łańcuchy lekkie połączone wiązaniami disulfidowymi w podstawowy kształt litery „Y” (rysunek 1). Pień cząsteczki w kształcie litery Y, stały region dwóch łańcuchów ciężkich, obejmuje błonę komórkową komórki B. Dwa miejsca wiążące antygen, znajdujące się na zewnątrz komórki B, są zaangażowane w wiązanie specyficznych epitopów patogenów w celu zainicjowania procesu aktywacji. Szacuje się, że każda naiwna dojrzała komórka B ma na swojej błonie do 100 000 BCR, a każdy z tych BCR ma identyczną swoistość wiązania epitopu.
Aby być przygotowanym do reakcji na szeroki zakres epitopów drobnoustrojów, komórki B, podobnie jak limfocyty T, wykorzystują genetyczną rearanżację setek segmentów genów w celu zapewnienia niezbędnej różnorodności swoistości receptorów. Zmienny region łańcucha ciężkiego BCR składa się z segmentów V, D i J, podobnie jak łańcuch β TCR. Region zmienny łańcucha lekkiego BCR składa się z segmentów V i J, podobnych do łańcucha α TCR. Genetyczna rearanżacja wszystkich możliwych kombinacji V-J-D (łańcuch ciężki) i V-J (łańcuch lekki) zapewnia miliony unikalnych miejsc wiążących antygen dla BCR i dla przeciwciał wydzielanych po aktywacji.
Jedną z istotnych różnic między BCR a TCR jest sposób, w jaki mogą one oddziaływać z epitopami antygenowymi. Podczas gdy TCR mogą oddziaływać tylko z epitopami antygenowymi, które są prezentowane w obrębie szczeliny wiążącej antygen w MHC I lub MHC II, BCR nie wymagają prezentacji antygenu za pomocą MHC; mogą oddziaływać z epitopami na wolnych antygenach lub z epitopami widocznymi na powierzchni nienaruszonych patogenów. Inną ważną różnicą jest to, że TCR rozpoznają tylko epitopy białkowe, podczas gdy BCR mogą rozpoznawać epitopy związane z różnymi klasami molekularnymi (np. białka, polisacharydy, lipopolisacharydy).
Aktywacja komórek B zachodzi poprzez różne mechanizmy w zależności od klasy molekularnej antygenu. Aktywacja komórki B przez antygen białkowy wymaga, aby komórka B funkcjonowała jako APC, prezentując epitopy białkowe za pomocą MHC II limfocytom T pomocniczym. Ze względu na ich zależność od komórek T w aktywacji komórek B, antygeny białkowe są klasyfikowane jako antygeny T-zależne. W przeciwieństwie do nich, polisacharydy, lipopolisacharydy i inne antygeny niebiałkowe są uważane za antygeny niezależne od T, ponieważ mogą aktywować komórki B bez przetwarzania i prezentacji antygenu komórkom T.
Pomyśl o tym
- Jakie typy cząsteczek służą jako BCR?
- Jakie są różnice między TCR i BCR w odniesieniu do rozpoznawania antygenów?
- Które klasy cząsteczek są antygenami T-zależnymi, a które antygenami T-niezależnymi?
Niezależna od komórek T aktywacja komórek B
Aktywacja komórek B bez współpracy pomocniczych komórek T jest określana jako aktywacja niezależna od komórek T i występuje, gdy BCR oddziałują z antygenami T-niezależnymi. Antygeny T-niezależne (np. kapsułki polisacharydowe, lipopolisacharyd) mają w swojej strukturze powtarzające się jednostki epitopowe, a to powtórzenie umożliwia krzyżowe wiązanie wielu BCR, zapewniając pierwszy sygnał do aktywacji (rysunek 2). Ponieważ komórki T nie biorą w tym udziału, drugi sygnał musi pochodzić z innych źródeł, takich jak interakcje receptorów toll-podobnych z PAMP lub interakcje z czynnikami układu dopełniacza.
Po aktywacji komórki B ulegają klonalnej proliferacji, a komórki potomne różnicują się w komórki plazmatyczne. Komórki plazmatyczne są fabrykami przeciwciał, które wydzielają duże ich ilości. Po zróżnicowaniu zanikają powierzchniowe BCR, a komórka plazmatyczna wydziela pentameryczne cząsteczki IgM, które mają taką samą swoistość antygenową jak BCR (rysunek 2).
Odpowiedź niezależna od komórek T jest krótkotrwała i nie prowadzi do wytworzenia komórek B pamięci. Dlatego nie spowoduje wtórnej odpowiedzi na kolejne ekspozycje na antygeny T-niezależne.

Rysunek 2. Antygeny T-niezależne mają powtarzające się epitopy, które mogą indukować rozpoznanie i aktywację komórek B bez udziału komórek T. Drugi sygnał, taki jak interakcja TLRs z PAMPs (nie pokazano), jest również wymagany do aktywacji komórki B. Po aktywacji, komórka B proliferuje i różnicuje się w komórki plazmatyczne wydzielające przeciwciała.
Pomyśl o tym
- Jakie są dwa sygnały wymagane do niezależnej od limfocytów T aktywacji komórek B?
- Jaka jest funkcja komórki plazmatycznej?
Zależna od komórek T aktywacja komórek B

Rysunek 3. Kliknij, aby powiększyć obraz. W zależnej od komórek T aktywacji komórek B, komórka B rozpoznaje i internalizuje antygen i przedstawia go limfocytowi pomocniczemu T, który jest specyficzny dla tego samego antygenu. Komórka pomocnicza T wchodzi w interakcję z antygenem prezentowanym przez komórkę B, co powoduje aktywację komórki T i stymuluje uwalnianie cytokin, które następnie aktywują komórkę B. Aktywacja komórek B wyzwala proliferację i różnicowanie w komórki B i komórki plazmatyczne.
Zależna od komórek T aktywacja komórek B jest bardziej złożona niż aktywacja niezależna od komórek T, ale wynikająca z niej odpowiedź immunologiczna jest silniejsza i rozwija pamięć. Aktywacja zależna od komórek T może wystąpić albo w odpowiedzi na wolne antygeny białkowe, albo na antygeny białkowe związane z nienaruszonym patogenem. Interakcja pomiędzy BCR na naiwnej dojrzałej komórce B i wolnym antygenem białkowym stymuluje internalizację antygenu, podczas gdy interakcja z antygenami związanymi z nienaruszonym patogenem inicjuje ekstrakcję antygenu z patogenu przed internalizacją. Po internalizacji wewnątrz komórki B, antygen białkowy jest przetwarzany i prezentowany za pomocą MHC II. Prezentowany antygen jest następnie rozpoznawany przez limfocyty T pomocnicze specyficzne dla tego samego antygenu. TCR pomocniczej komórki T rozpoznaje obcy antygen, a cząsteczka CD4 komórki T oddziałuje z MHC II na komórce B. Koordynacja pomiędzy komórkami B i pomocniczymi komórkami T, które są specyficzne dla tego samego antygenu, jest określana jako rozpoznanie sprzężone.
Po aktywacji przez rozpoznanie sprzężone, komórki TH2 produkują i wydzielają cytokiny, które aktywują komórki B i powodują proliferację do klonalnych komórek potomnych. Po kilku rundach proliferacji, dodatkowe cytokiny dostarczone przez komórki TH2 stymulują różnicowanie aktywowanych klonów komórek B w komórki pamięci B, które szybko reagują na kolejne ekspozycje na ten sam epitop białkowy, oraz komórki plazmatyczne, które tracą swoje błonowe BCR i początkowo wydzielają pentameryczne IgM (rysunek 3).
Po początkowym wydzielaniu IgM, cytokiny wydzielane przez komórki TH2 stymulują komórki plazmatyczne do przejścia z produkcji IgM na produkcję IgG, IgA lub IgE. Proces ten, zwany przełączaniem klas lub przełączaniem izotypów, umożliwia komórkom plazmatycznym sklonowanym z tej samej aktywowanej komórki B wytwarzanie różnych klas przeciwciał o tej samej swoistości epitopowej. Przełączanie klas jest osiągane przez genetyczną rearanżację segmentów genów kodujących region stały, który określa klasę przeciwciała. Region zmienny nie ulega zmianie, więc nowa klasa przeciwciała zachowuje pierwotną swoistość epitopową.
Pomyśl o tym
- Jakie kroki są wymagane do zależnej od komórek T aktywacji komórek B?
- Co to jest przełączanie klas przeciwciał i dlaczego jest ważne?
Pierwotna i wtórna odpowiedź
Zależna od komórek T aktywacja komórek B odgrywa ważną rolę zarówno w pierwotnej, jak i wtórnej odpowiedzi związanej z odpornością adaptacyjną. Przy pierwszym kontakcie z antygenem białkowym następuje zależna od komórek T pierwotna odpowiedź przeciwciał. Początkowym etapem odpowiedzi pierwotnej jest okres opóźnienia, lub okres utajony, trwający około 10 dni, podczas którego w surowicy nie można wykryć żadnego przeciwciała. Ten okres opóźnienia jest czasem wymaganym dla wszystkich etapów odpowiedzi pierwotnej, w tym wiązania antygenu przez naiwne dojrzałe komórki B z BCR, przetwarzania i prezentacji antygenu, aktywacji pomocniczych komórek T, aktywacji komórek B i proliferacji klonalnej. Koniec okresu opóźnienia charakteryzuje się wzrostem poziomu IgM w surowicy, ponieważ komórki TH2 stymulują różnicowanie komórek B w komórki plazmatyczne. Poziom IgM osiąga swój szczyt około 14 dni po ekspozycji na antygen pierwotny; mniej więcej w tym samym czasie TH2 stymuluje zmianę klasy przeciwciał, a poziom IgM w surowicy zaczyna spadać. W międzyczasie poziom IgG wzrasta, aż osiągnie szczyt około trzech tygodni od odpowiedzi pierwotnej (rysunek 4).
Podczas odpowiedzi pierwotnej niektóre ze sklonowanych komórek B różnicują się w komórki B pamięci, zaprogramowane do reagowania na kolejne ekspozycje. Ta wtórna odpowiedź występuje szybciej i mocniej niż odpowiedź pierwotna. Okres opóźnienia jest zmniejszony do zaledwie kilku dni, a produkcja IgG jest znacznie wyższa niż obserwowana w przypadku odpowiedzi pierwotnej (rysunek 4). Ponadto, przeciwciała produkowane podczas odpowiedzi wtórnej są bardziej skuteczne i wiążą się z wyższym powinowactwem do docelowych epitopów. Komórki plazmatyczne wytwarzane podczas odpowiedzi wtórnej żyją dłużej niż te wytwarzane podczas odpowiedzi pierwotnej, więc poziomy swoistych przeciwciał pozostają podwyższone przez dłuższy czas.

Ryc. 4. W porównaniu z odpowiedzią pierwotną, wtórna odpowiedź przeciwciał występuje szybciej i wytwarza poziomy przeciwciał, które są wyższe i bardziej trwałe. Odpowiedź wtórna obejmuje głównie IgG.
Pomyśl o tym
- Jakie zdarzenia występują podczas okresu opóźnienia pierwotnej odpowiedzi przeciwciał?
- Dlaczego poziom przeciwciał pozostaje podwyższony dłużej podczas wtórnej odpowiedzi przeciwciał?
Kluczowe pojęcia i podsumowanie
- Limfocyty B lub komórki B wytwarzają przeciwciała biorące udział w odporności humoralnej. Komórki B są produkowane w szpiku kostnym, gdzie zachodzą początkowe etapy dojrzewania, i wędrują do śledziony, gdzie zachodzą końcowe etapy dojrzewania do naiwnych dojrzałych komórek B.
- Receptory komórek B (BCR) są związanymi z błoną monomerycznymi formami IgD i IgM, które wiążą specyficzne epitopy antygenów za pomocą swoich regionów wiążących antygen Fab. Różnorodność specyficzności wiązania antygenu jest tworzona przez genetyczną rearanżację segmentów V, D i J, podobną do mechanizmu stosowanego w przypadku różnorodności TCR.
- Antygeny białkowe są nazywane antygenami T-zależnymi, ponieważ mogą one aktywować komórki B tylko przy współpracy pomocniczych komórek T. Inne klasy cząsteczek nie wymagają współpracy komórek T i są nazywane antygenami T-niezależnymi.
- Aktywacja komórek B niezależna od komórek T obejmuje krzyżowe wiązanie BCR przez powtarzające się epitopy antygenów niebiałkowych. Charakteryzuje się ona wytwarzaniem IgM przez komórki plazmatyczne i nie wytwarza komórek B pamięci.
- Zależna od komórek T aktywacja komórek B obejmuje przetwarzanie i prezentację antygenów białkowych pomocniczym komórkom T, aktywację komórek B przez cytokiny wydzielane z aktywowanych komórek TH2 oraz komórki plazmatyczne, które wytwarzają różne klasy przeciwciał w wyniku przełączenia klas. Wytwarzane są również komórki B pamięci.
- Ponowna ekspozycja na antygeny T-zależne powoduje wtórną odpowiedź przeciwciał inicjowaną przez komórki B pamięci. Odpowiedź wtórna rozwija się szybciej i wytwarza wyższe i bardziej trwałe poziomy przeciwciał o wyższym powinowactwie do swoistego antygenu.
Wielokrotny wybór
Który z poniższych byłby antygenem T-zależnym?
- lipopolysaccharide
- glycolipid
- protein
- carbohydrate
Which of the following would be a BCR?
- CD4
- MHC II
- MHC I
- IgD
Which of the following does not occur during the lag period of the primary antibody response?
- activation of helper T cells
- class switching to IgG
- presentation of antigen with MHC II
- binding of antigen to BCRs
Fill in the Blank
________ antigens can stimulate B cells to become activated but require cytokine assistance delivered by helper T cells.
T-independent antigens can stimulate B cells to become activated and secrete antibodies without assistance from helper T cells. These antigens possess ________ antigenic epitopes that cross-link BCRs.
Think about It
A patient lacks the ability to make functioning T cells because of a genetic disorder. Would this patient’s B cells be able to produce antibodies in response to an infection? Explain your answer.