B. Replicarea ADN-ului la bacterii
În general, ADN-ul se replică prin desfacerea spiralei, separarea catenei prin ruperea legăturilor de hidrogen dintre catenele complementare și sinteza a două noi catene prin împerechere de baze complementare. Replicația începe într-un loc specific din ADN numit origine de replicare (oriC).
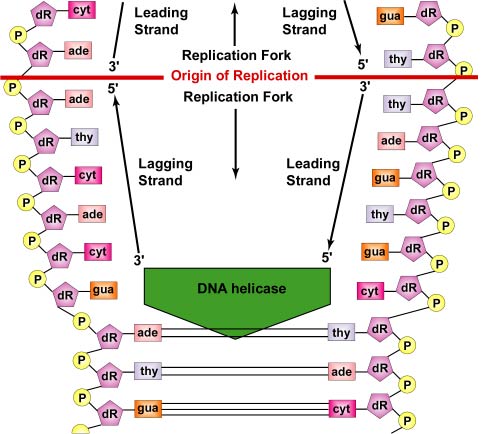
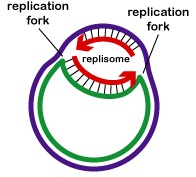
Replicarea ADN-ului este bidirecțională de la originea de replicare. Pentru a începe replicarea ADN, enzimele de derulare numite ADN-elicaze determină segmente scurte din cele două șiruri de ADN părinte să se deruleze și să se separe unul de celălalt la originea replicării pentru a forma două furci de replicare în formă de „Y”. Aceste furci de replicare reprezintă locul efectiv de copiere a ADN-ului (Figura \(\PageIndex{3}\)). Toate proteinele implicate în replicarea ADN-ului se agregă la furcile de replicare pentru a forma un complex de replicare numit replisom (Figura \(\PageIndex{4}\)).
Proteinele de legare a catenei simple se leagă de regiunile monocatenare, astfel încât cele două catene să nu se reunească. Desfășurarea helixului bicatenar generează superbobine pozitive în fața furcii de replicare. Enzimele numite topoizomeraze contracarează acest lucru prin producerea de rupturi în ADN și apoi le reunesc pentru a forma superbobine negative pentru a atenua acest stres în molecula elicoidală în timpul replicării.
În timp ce șuvițele continuă să se deruleze și să se separe în ambele direcții în jurul întregii molecule de ADN, noi șuvițe complementare sunt produse prin legarea de hidrogen a nucleotidelor libere de ADN cu cele de pe fiecare șuviță mamă. Pe măsură ce noile nucleotide se aliniază vizavi de fiecare filament părinte prin legătură de hidrogen, enzimele numite ADN polimeraze unesc nucleotidele prin intermediul legăturilor fosfodiester. De fapt, nucleotidele care se aliniază prin împerechere de baze complementare sunt trifosfați de deoxinucleotidă, compuși dintr-o bază azotată, deoxiriboză și trei fosfați. Pe măsură ce se formează legătura fosfodiester între grupul fosfat 5′ al noii nucleotide și OH 3′ al ultimei nucleotide din șirul de ADN, doi dintre fosfați sunt îndepărtați, furnizând energie pentru legare (a se vedea figura \(\PageIndex{6}\)). În final, fiecare catenă parentală servește drept șablon pentru a sintetiza o copie complementară a ei însăși, ceea ce duce la formarea a două molecule de ADN identice (a se vedea figura \(\PageIndex{7}\)). La bacterii, proteinele Par au rolul de a separa cromozomii bacterieni la polii opuși ai celulei în timpul diviziunii celulare. Ele se leagă de originea de replicare a ADN-ului și trag sau împing fizic cromozomii în afară, similar cu aparatul mitotic al celulelor eucariote. Proteinele Fts, cum ar fi FtsK din divizom, ajută, de asemenea, la separarea cromozomului bacterian replicat.
Animare GIF care ilustrează replicarea ADN-ului prin împerechere complementară de baze
În realitate, replicarea ADN-ului este mai complicată decât atât din cauza naturii ADN polimerazelor. Enzimele ADN-polimerazei sunt capabile doar să unească gruparea fosfat de la carbonul 5′ al unui nou nucleotid cu gruparea hidroxil (OH) de la carbonul 3′ al unui nucleotid aflat deja în lanț. Prin urmare, ADN-ul poate fi sintetizat numai în direcția 5′ – 3′, copiind un lanț părinte care se desfășoară în direcția 3′ – 5′.
Care șir de ADN are două capete. Capătul 5′ al ADN-ului este cel cu gruparea fosfat terminală pe carbonul 5′ al dezoxiribozei; capătul 3′ este cel cu o grupare hidroxil (OH) terminală pe dezoxiriboza carbonului 3′ al dezoxiribozei (a se vedea figura \(\PageIndex{8}\)). Cele două catene sunt antiparalele, adică merg în direcții opuse. Prin urmare, un singur șir părinte – cel care merge de la 3′ la 5′ și care se numește șir conducător – poate fi copiat direct pe toată lungimea sa (a se vedea figura \(\PageIndex{9}\)). Cu toate acestea, celălalt lanț părinte – cel care merge de la 5′ la 3′ și care se numește lanțul întârziat – trebuie copiat discontinuu în fragmente scurte (fragmente Okazaki) de aproximativ 100-1000 nucleotide fiecare, pe măsură ce ADN-ul se derulează. Acest lucru are loc, după cum s-a menționat mai sus, la nivelul replisomului. Șirul de ADN întârziat se desprinde în buclă de șirul conducător, ceea ce permite replisomului să se deplaseze de-a lungul ambelor șiruri, trăgând ADN-ul în timp ce are loc replicarea. ADN-ul propriu-zis, nu ADN polimeraza, este cel care se mișcă în timpul replicării ADN-ului bacterian (a se vedea figura \(\PageIndex{5}\)).
În plus, enzimele ADN polimerazei nu pot începe un nou lanț de ADN de la zero. Ele pot doar să atașeze noi nucleotide pe gruparea 3′ OH a unui nucleotid dintr-un lanț preexistent. Prin urmare, pentru a începe sinteza șirului conducător și a fiecărui fragment de ADN din șirul întârziat, este necesar un complex de ARN polimerază numit primază. Primaza, care este capabilă să unească nucleotide de ARN fără a avea nevoie de un catenar preexistent de acid nucleic, adaugă mai întâi câteva nucleotide de ARN comlementar opuse nucleotidelor de ADN de pe catena mamă. Acest lucru formează ceea ce se numește un primer de ARN (a se vedea figura \(\PageIndex{10}\\\)).
ADN polimeraza III înlocuiește apoi primaraza și este capabilă să adauge nucleotide de ADN la primerul de ARN (a se vedea figura \(\PageIndex{11}\)). Ulterior, ADN polimeraza II digeră amorța de ARN și înlocuiește nucleotidele de ARN ale amorței cu nucleotidele de ADN adecvate pentru a umple golul (a se vedea figura \(\PageIndex{12}\)). În cele din urmă, fragmentele de ADN sunt legate între ele de către enzima ADN ligază (a se vedea figura \(\PageIndex{9}\)). Cu toate acestea, chiar și cu această procedură complicată, o macromoleculă de 1000 de micrometri de lungime de ADN supraînfășurat și strâns împachetat poate realiza o copie exactă a sa în doar aproximativ 10 minute, în condiții optime, inserând nucleotide cu o rată de aproximativ 1000 de nucleotide pe secundă!
Animare a replicării ADN.
Courtesy of HHMI’s Biointeractive.