- Obiective de învățare
- Producția și maturarea celulelor B
- Gândiți-vă la asta
- Receptorii celulelor B
- Gândiți-vă
- Activarea independentă de celulele T a celulelor B
- Gândiți-vă
- Activarea dependentă de celulele T a celulelor B
- Gândiți-vă
- Răspunsuri primare și secundare
- Gândiți-vă la asta
- Concepte cheie și rezumat
- Multiple Choice
- Fill in the Blank
- Think about It
Obiective de învățare
- Descrieți producerea și maturizarea celulelor B
- Comparați structura receptorilor celulelor B și a receptorilor celulelor T
- Comparați receptorii celulelor T-activarea dependentă și T-independentă a celulelor B
- Compara răspunsurile primare și secundare ale anticorpilor
Imunitatea umană se referă la mecanismele de apărare imunitară adaptivă care sunt mediate de anticorpii secretați de limfocitele B, sau celule B. Această secțiune se va concentra asupra celulelor B și va discuta despre producția și maturarea lor, despre receptori și mecanismele de activare.
Producția și maturarea celulelor B
Ca și celulele T, celulele B se formează din celule stem hematopoietice multipotente (HSC) în măduva osoasă și urmează o cale prin celule stem limfoide și limfoblaste (a se vedea figura 1 din Apărări celulare). Totuși, spre deosebire de celulele T, limfoblastele destinate să devină celule B nu părăsesc măduva osoasă și nu se deplasează la timus pentru maturare. Mai degrabă, eventualele celule B continuă să se maturizeze în măduva osoasă.
Primul pas al maturării celulelor B este o evaluare a funcționalității receptorilor lor de legare a antigenului. Aceasta are loc prin selecția pozitivă pentru celulele B cu receptori funcționali normali. Un mecanism de selecție negativă este apoi utilizat pentru a elimina celulele B care reacționează singure și pentru a minimiza riscul de autoimunitate. Selecția negativă a celulelor B care reacționează singure poate implica eliminarea prin apoptoză, editarea sau modificarea receptorilor, astfel încât aceștia să nu mai fie autoreactivi, sau inducerea unei anergii în celulele B. Celulele B imature care trec de selecția din măduva osoasă se deplasează apoi în splină pentru etapele finale de maturare. Acolo ele devin celule B mature naive, adică, celule B mature care nu au fost încă activate.
Gândiți-vă la asta
- Comparați maturarea celulelor B cu maturarea celulelor T.
Receptorii celulelor B
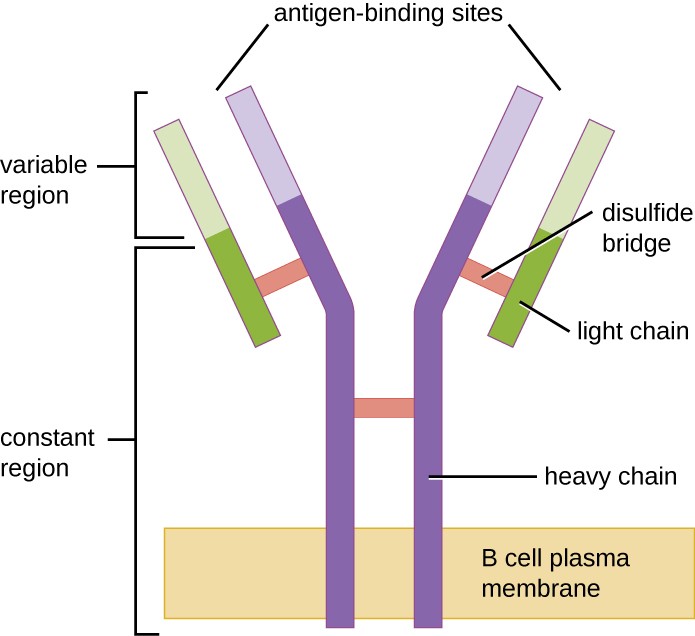
Figura 1. Receptorii celulelor B sunt încorporați în membranele celulelor B. Regiunile variabile ale tuturor receptorilor de pe o singură celulă se leagă de același antigen specific.
Ca și celulele T, celulele B posedă receptori specifici antigenului cu diverse specificități. Deși se bazează pe celulele T pentru o funcționare optimă, celulele B pot fi activate fără ajutor din partea celulelor T. Receptorii celulelor B (BCR) pentru celulele B mature naive sunt forme monomerice de IgD și IgM legate de membrană. Aceștia au două lanțuri grele identice și două lanțuri ușoare identice conectate prin legături disulfidice într-o formă de bază în formă de „Y” (figura 1). Trunchiul moleculei în formă de Y, regiunea constantă a celor două lanțuri grele, se întinde pe membrana celulelor B. Cele două situsuri de legare a antigenului expuse la exteriorul celulei B sunt implicate în legarea epitopilor patogeni specifici pentru a iniția procesul de activare. Se estimează că fiecare celulă B naivă matură matură are peste 100.000 de BCR-uri pe membrana sa, iar fiecare dintre aceste BCR-uri are o specificitate identică de legare a epitopilor.
Pentru a fi pregătite să reacționeze la o gamă largă de epitopi microbieni, celulele B, ca și celulele T, utilizează rearanjarea genetică a sute de segmente de gene pentru a asigura diversitatea necesară de specificități ale receptorilor. Regiunea variabilă a lanțului greu al BCR este alcătuită din segmentele V, D și J, similar cu lanțul β al TCR. Regiunea variabilă a lanțului ușor al BCR este alcătuită din segmentele V și J, asemănătoare lanțului α al TCR. Rearanjarea genetică a tuturor combinațiilor posibile de V-J-D (lanțul greu) și V-J (lanțul ușor) asigură milioane de situsuri unice de legare a antigenului pentru BCR și pentru anticorpii secretați după activare.
O diferență importantă între BCR și TCR este modul în care acestea pot interacționa cu epitopi antigenici. În timp ce TCR-urile pot interacționa numai cu epitopi antigenici care sunt prezentați în fanta de legare a antigenului din MHC I sau MHC II, BCR-urile nu au nevoie de prezentarea antigenului cu MHC; ele pot interacționa cu epitopi de pe antigene libere sau cu epitopi prezentați pe suprafața agenților patogeni intacți. O altă diferență importantă este că TCR-urile recunosc doar epitopi proteici, în timp ce BCR-urile pot recunoaște epitopi asociați cu diferite clase moleculare (de exemplu, proteine, polizaharide, lipopolizaharide).
Activarea celulelor B are loc prin diferite mecanisme, în funcție de clasa moleculară a antigenului. Activarea unei celule B de către un antigen proteic necesită ca celula B să funcționeze ca o APC, prezentând epitopii proteici cu MHC II către celulele T ajutătoare. Din cauza dependenței lor de celulele T pentru activarea celulelor B, antigenele proteice sunt clasificate ca antigene T-dependente. În schimb, polizaharidele, lipopolizaharidele și alte antigene neproteice sunt considerate antigene T-independente, deoarece pot activa celulele B fără procesarea și prezentarea antigenului la celulele T.
Gândiți-vă
- Ce tipuri de molecule servesc drept BCR?
- Care sunt diferențele dintre TCR și BCR în ceea ce privește recunoașterea antigenului?
- Ce clase de molecule sunt antigene dependente de T și care sunt antigene independente de T?
Activarea independentă de celulele T a celulelor B
Activarea celulelor B fără cooperarea celulelor T ajutătoare este denumită activare independentă de celulele T și are loc atunci când BCR-urile interacționează cu antigene independente de T. Antigenele T-independente (de exemplu, capsulele polizaharidice, lipopolizaharida) au în structura lor unități epitope repetitive, iar această repetiție permite încrucișarea mai multor BCR-uri, furnizând primul semnal de activare (figura 2). Deoarece celulele T nu sunt implicate, al doilea semnal trebuie să provină din alte surse, cum ar fi interacțiunile receptorilor toll-like cu PAMP-urile sau interacțiunile cu factorii din sistemul complementului.
După ce o celulă B este activată, aceasta suferă o proliferare clonală, iar celulele fiice se diferențiază în plasmocite. Plasmocitele sunt fabrici de anticorpi care secretă cantități mari de anticorpi. După diferențiere, BCR-urile de suprafață dispar și plasmocitele secretă molecule IgM pentamerice care au aceeași specificitate antigenică ca și BCR-urile (figura 2).
Răspunsul independent de celulele T este de scurtă durată și nu duce la producerea de celule B de memorie. Astfel, nu va avea ca rezultat un răspuns secundar la expuneri ulterioare la antigene T-independente.

Figura 2. Antigenele T-independente au epitopi repetitivi care pot induce recunoașterea și activarea celulei B fără implicarea celulelor T. Un al doilea semnal, cum ar fi interacțiunea TLR-urilor cu PAMP-urile (nu este prezentat), este, de asemenea, necesar pentru activarea celulei B. Odată activate, celulele B proliferează și se diferențiază în plasmocite secretoare de anticorpi.
Gândiți-vă
- Care sunt cele două semnale necesare pentru activarea independentă de celulele T a celulelor B?
- Care este funcția unei celule plasmatice?
Activarea dependentă de celulele T a celulelor B

Figura 3. Faceți clic pentru o imagine mai mare. În activarea dependentă de celulele T a celulelor B, celula B recunoaște și internalizează un antigen și îl prezintă unei celule T ajutătoare care este specifică aceluiași antigen. Celula T ajutătoare interacționează cu antigenul prezentat de celula B, care activează celula T și stimulează eliberarea de citokine care apoi activează celula B. Activarea celulei B declanșează proliferarea și diferențierea în celule B și plasmocite.
Activarea dependentă de celula T a celulelor B este mai complexă decât activarea independentă de celula T, dar răspunsul imunitar rezultat este mai puternic și dezvoltă memorie. Activarea dependentă de celulele T poate avea loc fie ca răspuns la antigene proteice libere, fie la antigene proteice asociate cu un agent patogen intact. Interacțiunea dintre BCR-urile de pe o celulă B naivă matură și un antigen proteic liber stimulează internalizarea antigenului, în timp ce interacțiunea cu antigene asociate cu un agent patogen intact inițiază extragerea antigenului din agentul patogen înainte de internalizare. Odată internalizat în interiorul celulei B, antigenul proteic este procesat și prezentat cu MHC II. Antigenul prezentat este apoi recunoscut de către celulele T ajutătoare specifice aceluiași antigen. TCR-ul celulei T ajutătoare recunoaște antigenul străin, iar molecula CD4 a celulei T interacționează cu MHC II de pe celula B. Coordonarea dintre celulele B și celulele T ajutătoare care sunt specifice aceluiași antigen se numește recunoaștere legată.
După ce sunt activate prin recunoașterea legată, celulele TH2 produc și secretă citokine care activează celula B și determină proliferarea în celule fiice clonale. După mai multe runde de proliferare, citokine suplimentare furnizate de celulele TH2 stimulează diferențierea clonelor de celule B activate în celule B de memorie, care vor răspunde rapid la expuneri ulterioare la același epitop proteic, și în celule plasmatice care își pierd BCR-urile de membrană și secretă inițial IgM pentamerice (figura 3).
După secreția inițială de IgM, citokinele secretate de celulele TH2 stimulează plasmocitele să treacă de la producția de IgM la producția de IgG, IgA sau IgE. Acest proces, numit comutare de clasă sau comutare de izotip, permite plasmocitelor clonate din aceeași celulă B activată să producă o varietate de clase de anticorpi cu aceeași specificitate de epitopi. Schimbarea clasei se realizează prin rearanjarea genetică a segmentelor de gene care codifică regiunea constantă, care determină clasa unui anticorp. Regiunea variabilă nu se modifică, astfel încât noua clasă de anticorpi își păstrează specificitatea epitopică inițială.
Gândiți-vă
- Ce etape sunt necesare pentru activarea dependentă de celulele T a celulelor B?
- Ce este schimbarea clasei de anticorpi și de ce este importantă?
Răspunsuri primare și secundare
Activarea dependentă de celulele T a celulelor B joacă un rol important atât în răspunsurile primare, cât și în cele secundare asociate cu imunitatea adaptativă. La prima expunere la un antigen proteic, are loc un răspuns primar al anticorpilor dependent de celulele T. Stadiul inițial al răspunsului primar este o perioadă de decalaj, sau perioadă latentă, de aproximativ 10 zile, în timpul căreia niciun anticorp nu poate fi detectat în ser. Această perioadă de decalaj este timpul necesar pentru toate etapele răspunsului primar, inclusiv legarea naivă a celulelor B mature de antigen cu BCR, procesarea și prezentarea antigenului, activarea celulelor T auxiliare, activarea celulelor B și proliferarea clonală. Sfârșitul perioadei de decalaj este caracterizat de o creștere a nivelului de IgM în ser, deoarece celulele TH2 stimulează diferențierea celulelor B în plasmocite. Nivelurile de IgM ating vârful în jur de 14 zile după expunerea primară la antigen; aproximativ în același timp, TH2 stimulează schimbarea clasei de anticorpi, iar nivelurile de IgM din ser încep să scadă. Între timp, nivelurile de IgG cresc până când ating un vârf după aproximativ trei săptămâni de răspuns primar (figura 4).
În timpul răspunsului primar, unele dintre celulele B clonate se diferențiază în celule B de memorie programate să răspundă la expuneri ulterioare. Acest răspuns secundar apare mai rapid și mai energic decât răspunsul primar. Perioada de decalaj este redusă la doar câteva zile, iar producția de IgG este semnificativ mai mare decât cea observată pentru răspunsul primar (figura 4). În plus, anticorpii produși în timpul răspunsului secundar sunt mai eficienți și se leagă cu o afinitate mai mare de epitopii vizați. Celulele plasmatice produse în timpul răspunsurilor secundare trăiesc mai mult decât cele produse în timpul răspunsului primar, astfel încât nivelurile de anticorpi specifici rămân ridicate pentru o perioadă mai lungă de timp.

Figura 4. În comparație cu răspunsul primar, răspunsul secundar al anticorpilor apare mai rapid și produce niveluri de anticorpi care sunt mai ridicate și mai susținute. Răspunsul secundar implică în principal IgG.
Gândiți-vă la asta
- Ce evenimente au loc în timpul perioadei de decalaj a răspunsului primar al anticorpilor?
- De ce nivelurile de anticorpi rămân ridicate mai mult timp în timpul răspunsului secundar al anticorpilor?
Concepte cheie și rezumat
- Limfocitele B sau celulele B produc anticorpi implicați în imunitatea umorală. Celulele B sunt produse în măduva osoasă, unde au loc etapele inițiale de maturare, și se deplasează la splină pentru etapele finale de maturare în celule B mature naive.
- Receptorii celulelor B (BCR) sunt forme monomerice de IgD și IgM legate de membrană care se leagă de epitopi antigeni specifici cu regiunile lor de legare a antigenului Fab. Diversitatea specificității de legare a antigenului este creată prin rearanjarea genetică a segmentelor V, D și J, similar mecanismului utilizat pentru diversitatea TCR.
- Antigenele proteice sunt numite antigene T-dependente, deoarece acestea pot activa celulele B numai cu cooperarea celulelor T ajutătoare. Alte clase de molecule nu necesită cooperarea celulelor T și se numesc antigene T-independente.
- Activarea independentă de celulele T a celulelor B implică încrucișarea BCR-urilor prin epitopi antigeni neproteici repetitivi. Se caracterizează prin producerea de IgM de către plasmocite și nu produce celule B cu memorie.
- Activarea dependentă de celulele T a celulelor B implică procesarea și prezentarea antigenelor proteice către celulele T ajutătoare, activarea celulelor B de către citokine secretate de celulele TH2 activate și plasmocite care produc diferite clase de anticorpi ca urmare a schimbării de clasă. Se produc, de asemenea, celule B de memorie.
- Expunerile secundare la antigeni T-dependenți au ca rezultat un răspuns secundar al anticorpilor inițiat de celulele B de memorie. Răspunsul secundar se dezvoltă mai rapid și produce niveluri mai ridicate și mai susținute de anticorpi cu afinitate mai mare pentru antigenul specific.
Multiple Choice
Care dintre următoarele ar fi un antigen T-dependent?
- lipopolysaccharide
- glycolipid
- protein
- carbohydrate
Which of the following would be a BCR?
- CD4
- MHC II
- MHC I
- IgD
Which of the following does not occur during the lag period of the primary antibody response?
- activation of helper T cells
- class switching to IgG
- presentation of antigen with MHC II
- binding of antigen to BCRs
Fill in the Blank
________ antigens can stimulate B cells to become activated but require cytokine assistance delivered by helper T cells.
T-independent antigens can stimulate B cells to become activated and secrete antibodies without assistance from helper T cells. These antigens possess ________ antigenic epitopes that cross-link BCRs.
Think about It
A patient lacks the ability to make functioning T cells because of a genetic disorder. Would this patient’s B cells be able to produce antibodies in response to an infection? Explain your answer.