B. Replikace DNA u bakterií
Obecně se DNA replikuje rozbalením šroubovice, oddělením vláken přerušením vodíkových vazeb mezi komplementárními vlákny a syntézou dvou nových vláken párováním komplementárních bází. Replikace začíná na určitém místě v DNA zvaném počátek replikace (oriC).
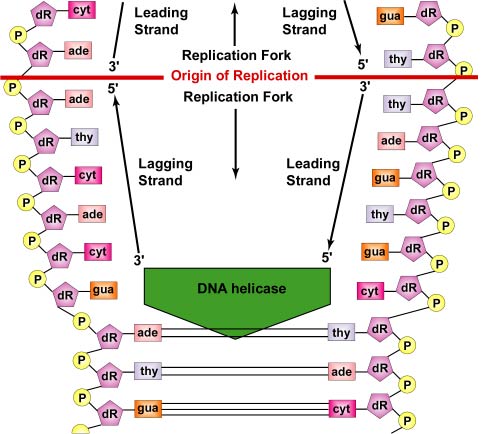
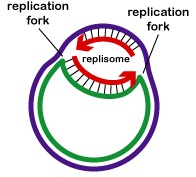
Replikace DNA je od počátku replikace obousměrná. Pro zahájení replikace DNA odvíjecí enzymy zvané DNA helikázy způsobí, že se krátké úseky obou mateřských vláken DNA odvíjejí a oddělují od sebe v místě počátku replikace a vytvářejí dvě replikační vidlice ve tvaru písmene „Y“. Tyto replikační vidlice jsou vlastním místem kopírování DNA (obrázek \(\PageIndex{3}\)). Všechny proteiny, které se podílejí na replikaci DNA, se shlukují v replikačních vidlicích a tvoří replikační komplex zvaný replisom (obrázek \(\PageIndex{4}\)).
Bílkoviny vázající jednořetězce se vážou na jednořetězcové oblasti, aby se obě vlákna znovu nespojila. Odvíjení dvouřetězcové šroubovice vytváří pozitivní superzávity před replikační vidličkou. Enzymy nazývané topoizomerázy tomu čelí tím, že vytvářejí zlomy v DNA a poté je znovu spojují a vytvářejí negativní superzávity, aby zmírnily toto napětí ve šroubovici během replikace.
Jak se vlákna dále odvíjejí a oddělují v obou směrech kolem celé molekuly DNA, vznikají nová komplementární vlákna vodíkovou vazbou volných nukleotidů DNA s těmi na každém rodičovském vlákně. Jak se nové nukleotidy vodíkovou vazbou řadí naproti každému rodičovskému vláknu, enzymy zvané DNA polymerázy spojují nukleotidy pomocí fosfodiesterových vazeb. Nukleotidy seřazené komplementárním párováním bází jsou vlastně deoxynukleotidtrifosfáty, které se skládají z dusíkaté báze, deoxyribózy a tří fosfátů. Při vytváření fosfodiesterové vazby mezi 5′ fosfátovou skupinou nového nukleotidu a 3′ OH posledního nukleotidu v řetězci DNA se odstraní dva fosfáty, které poskytují energii pro vazbu (viz obrázek \(\PageIndex{6}\)). Nakonec každé mateřské vlákno slouží jako šablona pro syntézu své komplementární kopie, čímž vznikají dvě identické molekuly DNA (viz obrázek \(\PageIndex{7}\)). V bakteriích slouží proteiny Par k oddělení bakteriálních chromozomů na opačných pólech buňky během buněčného dělení. Vážou se na počátek replikace DNA a fyzicky táhnou nebo tlačí chromozomy od sebe, podobně jako mitotický aparát eukaryotických buněk. Při oddělování replikovaného bakteriálního chromozomu pomáhají také proteiny Fts, například FtsK v divizomu.
Animace GIF znázorňující replikaci DNA pomocí komplementárního párování bází
Ve skutečnosti je replikace DNA složitější, a to kvůli povaze polymeráz DNA. Enzymy DNA polymerázy jsou schopny pouze připojit fosfátovou skupinu na 5′ uhlíku nového nukleotidu k hydroxylové (OH) skupině na 3′ uhlíku nukleotidu, který již v řetězci je. Výsledkem je, že DNA lze syntetizovat pouze ve směru 5′ až 3′ při kopírování mateřského řetězce probíhajícího ve směru 3′ až 5′.
Každé vlákno DNA má dva konce. Konec 5′ DNA je ten s terminální fosfátovou skupinou na 5′ uhlíku deoxyribózy; konec 3′ je ten s terminální hydroxylovou (OH) skupinou na 3′ uhlíku deoxyribózy (viz obrázek \(\PageIndex{8}\). Obě vlákna jsou antiparalelní, to znamená, že probíhají v opačných směrech. Proto lze jedno mateřské vlákno – to, které vede od 3′ k 5′ a nazývá se vedoucí vlákno – kopírovat přímo po celé jeho délce (viz obrázek \(\PageIndex{9}\)). Druhé mateřské vlákno, které vede od 5′ ke 3′ a nazývá se opožděné vlákno, se však musí kopírovat přerušovaně v krátkých fragmentech (Okazakiho fragmenty) po přibližně 100-1000 nukleotidech, jak se DNA odvíjí. K tomu dochází, jak bylo uvedeno výše, v replisomu. Zpožděné vlákno DNA se odpojuje od vedoucího vlákna, což umožňuje replisomu pohybovat se podél obou vláken a protahovat DNA při replikaci. Při replikaci bakteriální DNA se pohybuje vlastní DNA, nikoli DNA polymeráza (viz obrázek \(\PageIndex{5}\)).
Enzymy DNA polymerázy navíc nemohou začít nový řetězec DNA od nuly. Mohou pouze připojit nové nukleotidy na 3′ OH skupinu nukleotidu v již existujícím řetězci. Proto je k zahájení syntézy vedoucího řetězce a každého fragmentu DNA zaostávajícího řetězce zapotřebí komplex RNA polymerázy nazývaný primáza. Primáza, která je schopna spojovat nukleotidy RNA, aniž by vyžadovala již existující vlákno nukleové kyseliny, nejprve přidá několik komplementárních nukleotidů RNA naproti nukleotidům DNA na rodičovském vlákně. Tím vznikne tzv. RNA primer (viz obrázek \(\PageIndex{10}\).
DNA polymeráza III poté nahradí primázu a je schopna přidat nukleotidy DNA k RNA primeru (viz obrázek \(\PageIndex{11}\). Později DNA polymeráza II odštěpí RNA primer a nahradí nukleotidy RNA primeru správnými nukleotidy DNA, aby vyplnila mezeru (viz obrázek \(\PageIndex{12}\)). Nakonec jsou samotné fragmenty DNA spojeny enzymem DNA ligázou (viz obrázek \(\PageIndex{9}\)). I při tomto složitém postupu však může 1000 mikrometrů dlouhá makromolekula pevně zabalené, superzavinuté DNA za optimálních podmínek vytvořit přesnou kopii sebe sama za pouhých 10 minut, přičemž vkládá nukleotidy rychlostí asi 1000 nukleotidů za sekundu!
Animace replikace DNA.
Díky společnosti HHMI’s Biointeractive.