- Objectifs d’apprentissage
- Production et maturation des cellules B
- Réfléchissez-y
- Récepteurs des cellules B
- Réfléchissez-y
- Activation des cellules B indépendante des cellules T
- Réfléchissez-y
- Activation des lymphocytes B dépendante des cellules T
- Réfléchissez-y
- Réponses primaire et secondaire
- Réfléchissez-y
- Concepts clés et résumé
- Choix multiple
- Fill in the Blank
- Think about It
Objectifs d’apprentissage
- Décrire la production et la maturation des cellules B
- Comparer la structure des récepteurs des cellules B et des récepteurs des cellules T
- Comparer l’activation des cellules B dépendante et indépendante de T-.dépendante et T-indépendante des cellules B
- Comparer les réponses anticorps primaires et secondaires
L’immunité humorale fait référence aux mécanismes des défenses immunitaires adaptatives qui sont médiés par les anticorps sécrétés par les lymphocytes B, ou cellules B. Cette section se concentrera sur les cellules B et abordera leur production et leur maturation, leurs récepteurs et leurs mécanismes d’activation.
Production et maturation des cellules B
Comme les cellules T, les cellules B sont formées à partir de cellules souches hématopoïétiques (CSH) multipotentes dans la moelle osseuse et suivent une voie passant par la cellule souche lymphoïde et le lymphoblaste (voir la figure 1 dans les défenses cellulaires). Cependant, contrairement aux cellules T, les lymphoblastes destinés à devenir des cellules B ne quittent pas la moelle osseuse et ne se rendent pas dans le thymus pour leur maturation. Au contraire, les éventuels lymphocytes B continuent à mûrir dans la moelle osseuse.
La première étape de la maturation des lymphocytes B est une évaluation de la fonctionnalité de leurs récepteurs de liaison à l’antigène. Cela se fait par une sélection positive des cellules B ayant des récepteurs fonctionnels normaux. Un mécanisme de sélection négative est ensuite utilisé pour éliminer les cellules B autoréactives et minimiser le risque d’auto-immunité. La sélection négative des cellules B autoréactives peut impliquer l’élimination par apoptose, l’édition ou la modification des récepteurs afin qu’ils ne soient plus autoréactifs, ou l’induction d’une anergie dans la cellule B. Les cellules B immatures qui passent la sélection dans la moelle osseuse se déplacent ensuite vers la rate pour les dernières étapes de leur maturation. Là, ils deviennent des lymphocytes B matures naïfs, c’est-à-dire, des cellules B matures qui n’ont pas encore été activées.
Réfléchissez-y
- Comparer la maturation des cellules B à celle des cellules T.
Récepteurs des cellules B
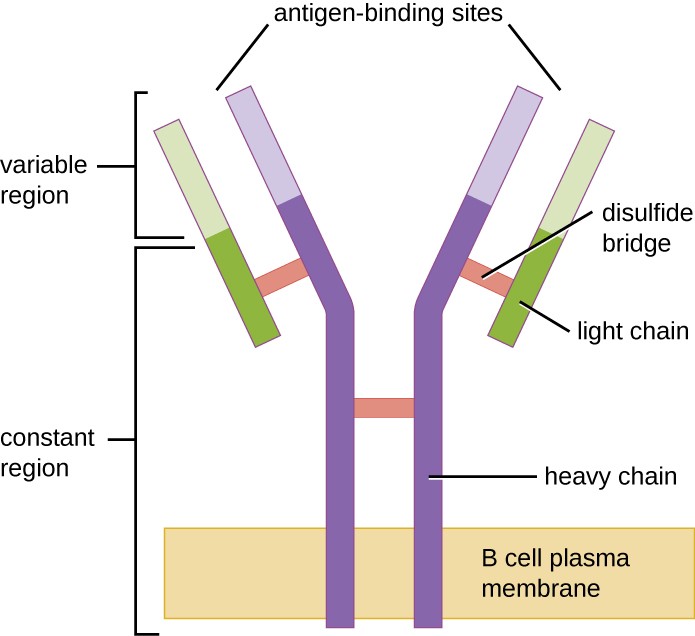
Figure 1. Les récepteurs des cellules B sont intégrés dans les membranes des cellules B. Les régions variables de tous les récepteurs d’une même cellule se lient au même antigène spécifique.
Comme les cellules T, les cellules B possèdent des récepteurs spécifiques de l’antigène avec des spécificités diverses. Bien qu’ils dépendent des cellules T pour un fonctionnement optimal, les cellules B peuvent être activées sans l’aide des cellules T. Les récepteurs des cellules B (BCR) des cellules B matures naïves sont des formes monomères d’IgD et d’IgM liées à la membrane. Ils possèdent deux chaînes lourdes identiques et deux chaînes légères identiques reliées par des liaisons disulfure en forme de « Y » de base (figure 1). Le tronc de la molécule en forme de Y, la région constante des deux chaînes lourdes, enjambe la membrane du lymphocyte B. Les deux sites de liaison à l’antigène exposés à l’extérieur de la cellule B sont impliqués dans la liaison d’épitopes pathogènes spécifiques pour initier le processus d’activation. On estime que chaque cellule B mature naïve possède plus de 100 000 BCR sur sa membrane, et que chacun de ces BCR a une spécificité de liaison aux épitopes identique.
Afin d’être préparés à réagir à une large gamme d’épitopes microbiens, les cellules B, comme les cellules T, utilisent le réarrangement génétique de centaines de segments de gènes pour fournir la diversité nécessaire des spécificités des récepteurs. La région variable de la chaîne lourde du BCR est constituée des segments V, D et J, comme la chaîne β du TCR. La région variable de la chaîne légère du BCR est constituée de segments V et J, similaires à la chaîne α du TCR. Le réarrangement génétique de toutes les combinaisons possibles de V-J-D (chaîne lourde) et V-J (chaîne légère) permet d’obtenir des millions de sites uniques de liaison à l’antigène pour le BCR et pour les anticorps sécrétés après activation.
Une différence importante entre les BCR et les TCR est la manière dont ils peuvent interagir avec les épitopes antigéniques. Alors que les TCR ne peuvent interagir qu’avec des épitopes antigéniques présentés dans la fente de liaison à l’antigène du CMH I ou du CMH II, les BCR ne nécessitent pas de présentation de l’antigène avec le CMH ; ils peuvent interagir avec des épitopes sur des antigènes libres ou avec des épitopes affichés à la surface d’agents pathogènes intacts. Une autre différence importante est que les TCR ne reconnaissent que des épitopes protéiques, alors que les BCR peuvent reconnaître des épitopes associés à différentes classes moléculaires (par exemple, des protéines, des polysaccharides, des lipopolysaccharides).
L’activation des cellules B se produit par différents mécanismes selon la classe moléculaire de l’antigène. L’activation d’une cellule B par un antigène protéique nécessite que la cellule B fonctionne comme une APC, présentant les épitopes protéiques avec le CMH II aux cellules T auxiliaires. En raison de leur dépendance vis-à-vis des lymphocytes T pour l’activation des cellules B, les antigènes protéiques sont classés comme des antigènes T-dépendants. En revanche, les polysaccharides, les lipopolysaccharides et d’autres antigènes non protéiques sont considérés comme des antigènes T-indépendants, car ils peuvent activer les cellules B sans traitement et présentation de l’antigène aux cellules T.
Réfléchissez-y
- Quels types de molécules servent de BCR ?
- Quelles sont les différences entre les TCR et les BCR en ce qui concerne la reconnaissance des antigènes ?
- Quelles classes de molécules sont des antigènes T-dépendants et quelles sont les antigènes T-indépendants ?
Activation des cellules B indépendante des cellules T
L’activation des cellules B sans la coopération des cellules T auxiliaires est appelée activation indépendante des cellules T et se produit lorsque les BCR interagissent avec des antigènes T-indépendants. Les antigènes indépendants des cellules T (par exemple, les capsules de polysaccharides, le lipopolysaccharide) ont des unités épitopiques répétitives dans leur structure, et cette répétition permet la liaison croisée de plusieurs BCR, fournissant le premier signal d’activation (Figure 2). Les lymphocytes T n’étant pas impliqués, le second signal doit provenir d’autres sources, comme les interactions des récepteurs de type péage avec les PAMP ou les interactions avec les facteurs du système du complément.
Une fois qu’un lymphocyte B est activé, il subit une prolifération clonale et les cellules filles se différencient en plasmocytes. Les plasmocytes sont des usines à anticorps qui sécrètent de grandes quantités d’anticorps. Après différenciation, les BCR de surface disparaissent et le plasmocyte sécrète des molécules IgM pentamériques qui ont la même spécificité antigénique que les BCR (figure 2).
La réponse indépendante des lymphocytes T est de courte durée et n’entraîne pas la production de lymphocytes B mémoire. Elle n’entraînera donc pas de réponse secondaire lors d’expositions ultérieures à des antigènes indépendants des lymphocytes T.

Figure 2. Les antigènes indépendants des cellules T ont des épitopes répétitifs qui peuvent induire la reconnaissance et l’activation des cellules B sans implication des cellules T. Un deuxième signal, tel que l’interaction des TLR avec les PAMPs (non montré), est également nécessaire pour l’activation de la cellule B. Une fois activé, le lymphocyte B prolifère et se différencie en plasmocytes sécréteurs d’anticorps.
Réfléchissez-y
- Quels sont les deux signaux nécessaires à l’activation des lymphocytes B indépendamment des lymphocytes T ?
- Quelle est la fonction d’un plasmocyte ?
Activation des lymphocytes B dépendante des cellules T

Figure 3. Cliquez pour agrandir l’image. Dans l’activation des cellules B dépendante des cellules T, la cellule B reconnaît et internalise un antigène et le présente à une cellule T auxiliaire spécifique du même antigène. Le lymphocyte T auxiliaire interagit avec l’antigène présenté par le lymphocyte B, ce qui active le lymphocyte T et stimule la libération de cytokines qui activent ensuite le lymphocyte B. L’activation du lymphocyte B déclenche le déclenchement d’une réponse immunitaire. L’activation de la cellule B déclenche la prolifération et la différenciation en cellules B et en plasmocytes.
L’activation des cellules B dépendante des cellules T est plus complexe que l’activation indépendante des cellules T, mais la réponse immunitaire qui en résulte est plus forte et développe une mémoire. L’activation dépendante des lymphocytes T peut se produire soit en réponse à des antigènes protéiques libres, soit à des antigènes protéiques associés à un agent pathogène intact. L’interaction entre les BCR d’une cellule B mature naïve et un antigène protéique libre stimule l’internalisation de l’antigène, tandis que l’interaction avec des antigènes associés à un agent pathogène intact déclenche l’extraction de l’antigène de l’agent pathogène avant son internalisation. Une fois internalisé dans la cellule B, l’antigène protéique est traité et présenté par le CMH II. L’antigène présenté est ensuite reconnu par les cellules T auxiliaires spécifiques du même antigène. Le TCR de la cellule T auxiliaire reconnaît l’antigène étranger et la molécule CD4 de la cellule T interagit avec le CMH II sur la cellule B. La coordination entre les cellules B et les cellules T auxiliaires spécifiques du même antigène est appelée reconnaissance liée.
Une fois activées par la reconnaissance liée, les cellules TH2 produisent et sécrètent des cytokines qui activent la cellule B et provoquent une prolifération en cellules filles clonales. Après plusieurs cycles de prolifération, des cytokines supplémentaires fournies par les cellules TH2 stimulent la différenciation des clones de cellules B activées en cellules B à mémoire, qui répondront rapidement à des expositions ultérieures au même épitope protéique, et en plasmocytes qui perdent leurs BCR membranaires et sécrètent initialement des IgM pentamériques (figure 3).
Après la sécrétion initiale d’IgM, les cytokines sécrétées par les cellules TH2 stimulent les plasmocytes pour qu’ils passent de la production d’IgM à celle d’IgG, d’IgA ou d’IgE. Ce processus, appelé changement de classe ou changement d’isotype, permet aux plasmocytes clonés à partir de la même cellule B activée de produire diverses classes d’anticorps ayant la même spécificité épitopique. Le changement de classe s’effectue par réarrangement génétique des segments de gènes codant pour la région constante, qui détermine la classe d’un anticorps. La région variable n’est pas modifiée, de sorte que la nouvelle classe d’anticorps conserve la spécificité d’épitope initiale.
Réfléchissez-y
- Quelles sont les étapes nécessaires à l’activation des lymphocytes B par les lymphocytes T ?
- Qu’est-ce que le changement de classe des anticorps et pourquoi est-il important ?
Réponses primaire et secondaire
L’activation des cellules B dépendante des lymphocytes T joue un rôle important dans les réponses primaire et secondaire associées à l’immunité adaptative. Lors de la première exposition à un antigène protéique, une réponse anticorps primaire dépendante des lymphocytes T se produit. L’étape initiale de la réponse primaire est une période de latence, ou période de latence, d’environ 10 jours, pendant laquelle aucun anticorps ne peut être détecté dans le sérum. Cette période de latence est le temps nécessaire à toutes les étapes de la réponse primaire, y compris la liaison des cellules B matures naïves avec les BCR, le traitement et la présentation de l’antigène, l’activation des cellules T auxiliaires, l’activation des cellules B et la prolifération clonale. La fin de la période de latence est caractérisée par une augmentation des taux d’IgM dans le sérum, car les cellules TH2 stimulent la différenciation des cellules B en plasmocytes. Les taux d’IgM atteignent leur maximum environ 14 jours après l’exposition à l’antigène primaire ; à peu près au même moment, le TH2 stimule le changement de classe d’anticorps, et les taux d’IgM dans le sérum commencent à diminuer. Pendant ce temps, les taux d’IgG augmentent jusqu’à atteindre un pic environ trois semaines après la réponse primaire (figure 4).
Pendant la réponse primaire, certains des lymphocytes B clonés se différencient en lymphocytes B mémoire programmés pour répondre à des expositions ultérieures. Cette réponse secondaire se produit plus rapidement et plus fortement que la réponse primaire. La période de latence est réduite à quelques jours seulement et la production d’IgG est significativement plus élevée que celle observée pour la réponse primaire (Figure 4). En outre, les anticorps produits au cours de la réponse secondaire sont plus efficaces et se lient avec une plus grande affinité aux épitopes ciblés. Les plasmocytes produits au cours des réponses secondaires vivent plus longtemps que ceux produits au cours de la réponse primaire, de sorte que les niveaux d’anticorps spécifiques restent élevés pendant une période plus longue.

Figure 4. Par rapport à la réponse primaire, la réponse secondaire en anticorps se produit plus rapidement et produit des taux d’anticorps plus élevés et plus durables. La réponse secondaire implique principalement les IgG.
Réfléchissez-y
- Quels événements se produisent pendant la période de latence de la réponse anticorps primaire ?
- Pourquoi les taux d’anticorps restent-ils élevés plus longtemps pendant la réponse anticorps secondaire ?
Concepts clés et résumé
- Les lymphocytes B ou cellules B produisent des anticorps impliqués dans l’immunité humorale. Les cellules B sont produites dans la moelle osseuse, où se produisent les étapes initiales de la maturation, et se déplacent vers la rate pour les étapes finales de la maturation en cellules B matures naïves.
- Les récepteurs des cellules B (BCR) sont des formes monomères membranaires d’IgD et d’IgM qui se lient à des épitopes antigéniques spécifiques avec leurs régions Fab de liaison à l’antigène. La diversité de la spécificité de liaison à l’antigène est créée par un réarrangement génétique des segments V, D et J, similaire au mécanisme utilisé pour la diversité des TCR.
- Les antigènes protéiques sont appelés antigènes T-dépendants car ils ne peuvent activer les cellules B qu’avec la coopération des cellules T auxiliaires. D’autres classes de molécules ne nécessitent pas la coopération des cellules T et sont appelées antigènes indépendants des cellules T.
- L’activation des cellules B indépendante des cellules T implique une liaison croisée des BCR par des épitopes antigéniques non protéiques répétitifs. Elle se caractérise par la production d’IgM par les plasmocytes et ne produit pas de cellules B à mémoire.
- L’activation des cellules B dépendante des cellules T implique le traitement et la présentation d’antigènes protéiques aux cellules T auxiliaires, l’activation des cellules B par des cytokines sécrétées par les cellules TH2 activées, et des plasmocytes qui produisent différentes classes d’anticorps suite à un changement de classe. Des cellules B à mémoire sont également produites.
- Les expositions secondaires à des antigènes T-dépendants entraînent une réponse anticorps secondaire initiée par les cellules B à mémoire. La réponse secondaire se développe plus rapidement et produit des niveaux plus élevés et plus soutenus d’anticorps avec une affinité plus élevée pour l’antigène spécifique.
Choix multiple
Lequel des éléments suivants serait un antigène T-dépendant ?
- lipopolysaccharide
- glycolipid
- protein
- carbohydrate
Which of the following would be a BCR?
- CD4
- MHC II
- MHC I
- IgD
Which of the following does not occur during the lag period of the primary antibody response?
- activation of helper T cells
- class switching to IgG
- presentation of antigen with MHC II
- binding of antigen to BCRs
Fill in the Blank
________ antigens can stimulate B cells to become activated but require cytokine assistance delivered by helper T cells.
T-independent antigens can stimulate B cells to become activated and secrete antibodies without assistance from helper T cells. These antigens possess ________ antigenic epitopes that cross-link BCRs.
Think about It
A patient lacks the ability to make functioning T cells because of a genetic disorder. Would this patient’s B cells be able to produce antibodies in response to an infection? Explain your answer.