B. DNS-replikáció a baktériumokban
A DNS replikációja általában a spirál feltekeredésével, a komplementer szálak közötti hidrogénkötések felbontásával történő szálleválasztással és a két új szál komplementer bázispárosodással történő szintézisével történik. A replikáció a DNS egy meghatározott helyén, a replikációs origónál (oriC) kezdődik.
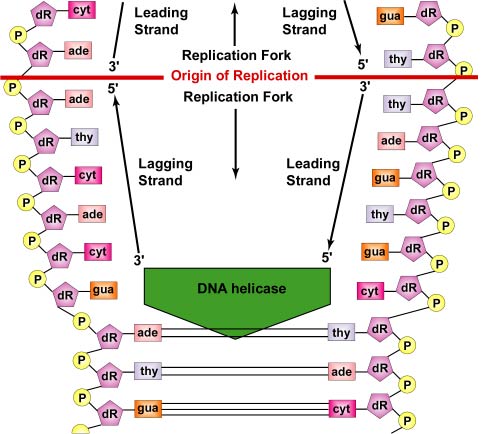
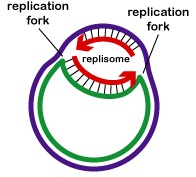
A DNS-replikáció a replikációs origótól kétirányú. A DNS-replikáció megkezdéséhez a DNS-helikázoknak nevezett kitekerő enzimek hatására a két szülő DNS-szál rövid szakaszai kitekerednek és elválnak egymástól a replikáció origójánál, hogy két “Y” alakú replikációs villát alkossanak. Ezek a replikációs villák a DNS-másolás tényleges helyszínei (\(\PageIndex{3}\) ábra). A DNS-replikációban részt vevő összes fehérje a replikációs villáknál egy replikációs komplexet, a repliszómát alkotja (\(\(\PageIndex{4}\) ábra).
Az egyszálas kötőfehérjék kötődnek az egyszálas régiókhoz, így a két szál nem kapcsolódik újra. A kettős szálú spirál kitekeredése pozitív szupertekercseket hoz létre a replikációs villa előtt. A topoizomerázoknak nevezett enzimek ezt úgy ellensúlyozzák, hogy töréseket hoznak létre a DNS-ben, majd újra egyesítik azokat, hogy negatív szupertekercseket képezzenek, hogy a replikáció során enyhítsék ezt a feszültséget a spirálmolekulában.
Miközben a szálak tovább tekerednek és szétválnak mindkét irányban a teljes DNS-molekula körül, új komplementer szálak keletkeznek a szabad DNS-nukleotidok hidrogénkötése révén az egyes szülői szálakhoz. Ahogy az új nukleotidok a hidrogénkötés révén felsorakoznak az egyes szülői szálakkal szemben, a DNS-polimerázoknak nevezett enzimek foszfodiészterkötések révén összekapcsolják a nukleotidokat. A komplementer bázispárosodás révén felsorakozó nukleotidok valójában dezoxinukleotid-trifoszfátok, amelyek egy nitrogénbázisból, dezoxiribózból és három foszfátból állnak. Ahogy a foszfodiészterkötés kialakul az új nukleotid 5′ foszfátcsoportja és a DNS-szál utolsó nukleotidjának 3′ OH-csoportja között, a foszfátok közül kettő eltávolodik, ami energiát biztosít a kötéshez (lásd \(\PageIndex{6}\) ábra). Végül minden szülői szál sablonként szolgál saját maga komplementer példányának szintéziséhez, így két azonos DNS-molekula jön létre (lásd \(\(\PageIndex{7}\) ábra). A baktériumokban a Par-fehérjék feladata, hogy a sejtosztódás során a bakteriális kromoszómákat a sejt ellentétes pólusaira szétválasszák. A DNS replikációs origójához kötődnek, és fizikailag széthúzzák vagy szétnyomják a kromoszómákat, hasonlóan az eukarióta sejtek mitotikus apparátusához. Az Fts-fehérjék, mint például a diviszómában található FtsK, szintén segítenek a replikált bakteriális kromoszóma szétválasztásában.
GIF animáció, amely a DNS replikációt illusztrálja a komplementer bázispárosítással
A valóságban a DNS replikáció ennél bonyolultabb a DNS-polimerázok természete miatt. A DNS-polimeráz enzimek csak arra képesek, hogy egy új nukleotid 5′ szénatomjánál lévő foszfátcsoportot egy már a láncban lévő nukleotid 3′ szénatomjának hidroxil (OH) csoportjához kapcsolják. Ennek eredményeképpen a DNS csak 5′ – 3′ irányban szintetizálható, miközben egy 3′ – 5′ irányban futó anyaszálat másolnak.
Minden DNS-szálnak két vége van. A DNS 5′ vége az, amelynek a dezoxiribóz 5′ szénatomján terminális foszfátcsoport található; a 3′ vége az, amelynek a dezoxiribóz 3′ szénatomján terminális hidroxil (OH) csoport található (lásd \(\PageIndex{8}\) ábra). A két szál antiparallel, azaz ellentétes irányban fut. Ezért az egyik szülői szál – a 3′-tól 5′-ig tartó, vezető szálnak nevezett – közvetlenül másolható a teljes hosszában (lásd \(\(\PageIndex{9}\) ábra). A másik szülői szálat azonban – az 5′-től 3′-ig futó szálat, amelyet lemaradó szálnak nevezünk – a DNS kitekeredése során rövid, egyenként körülbelül 100-1000 nukleotidból álló fragmentumokban (Okazaki-fragmentumok) diszkontinuusan kell lemásolni. Ez, mint már említettük, a replikoszómában történik. A lemaradó DNS-szál kilóg a vezető szálból, és ez lehetővé teszi, hogy a repliszóma mindkét szál mentén mozogjon, áthúzva a DNS-t a replikáció során. A bakteriális DNS-replikáció során nem a DNS-polimeráz, hanem a tényleges DNS mozog (lásd a \(\PageIndex{5}\) ábra).
A DNS-polimeráz enzimek ráadásul nem tudnak a semmiből új DNS-láncot kezdeni. Csak egy már meglévő szálban lévő nukleotid 3′ OH-csoportjához tudnak új nukleotidokat csatolni. Ezért a vezető szál és a lemaradó szál egyes DNS-töredékeinek szintézisének elindításához egy primáznak nevezett RNS-polimeráz-komplexre van szükség. A primáz, amely képes az RNS-nukleotidok összekapcsolására anélkül, hogy szükség lenne egy már létező nukleinsav-szálra, először több komplementer RNS-nukleotidot ad hozzá az alapszálon lévő DNS-nukleotidokkal szemben. Ezáltal egy úgynevezett RNS-primer alakul ki (lásd a \(\PageIndex{10}\) ábrát).
Azután a DNS-polimeráz III felváltja a primázt, és képes DNS-nukleotidokat hozzáadni az RNS-primerhez (lásd a \(\PageIndex{11}\) ábrát). Később a DNS-polimeráz II elemészti az RNS-primert, és a primer RNS-nukleotidjait a megfelelő DNS-nukleotidokkal helyettesíti, hogy kitöltse a rést (lásd a \(\PageIndex{12}\) ábrát). Végül magukat a DNS-darabkákat a DNS-ligáz enzim köti össze (lásd \(\(\PageIndex{9}\) ábra). Még ezzel a bonyolult eljárással együtt is, egy 1000 mikrométer hosszú, szorosan összecsomagolt, szupertekercselt DNS makromolekula optimális körülmények között mindössze körülbelül 10 perc alatt képes pontos másolatot készíteni magáról, másodpercenként körülbelül 1000 nukleotidot illesztve be!
A DNS replikáció animációja.
A HHMI’s Biointeractive jóvoltából.