- Tanulmányi célok
- B-sejtek termelődése és érése
- Gondolj bele
- B-sejt receptorok
- Gondolj bele
- A B-sejtek T-sejt-független aktivációja
- Gondolj bele
- A B-sejtek T-sejt-függő aktivációja
- Gondolj bele
- Primer és másodlagos válaszok
- Gondolj bele
- Főbb fogalmak és összefoglalás
- Multiple Choice
- Fill in the Blank
- Think about It
Tanulmányi célok
- Írd le a B-sejtek termelődését és érését
- Hasonlítsd össze a B-sejt receptorok és a T-sejt receptorok szerkezetét
- Hasonlítsd össze a T- és a T-sejt receptorok szerkezetét
- Hasonlítsd össze a T- és a T-sejt receptorok szerkezetét
- .B-sejtek T-függő és T-független aktivációját
- Hasonlítsa össze az elsődleges és másodlagos antitestválaszokat
A humorális immunitás az adaptív immunvédekezés azon mechanizmusaira utal, amelyeket a B-limfociták által szekretált antitestek közvetítenek, vagy B-sejtek által kibocsátott sejtek. Ez a szakasz a B-sejtekre összpontosít, és a termelésüket és érésüket, receptoraikat és aktivációs mechanizmusaikat tárgyalja.
B-sejtek termelődése és érése
A T-sejtekhez hasonlóan a B-sejtek is a csontvelőben lévő multipotens vérképző őssejtekből (HSC) képződnek, és a limfoid őssejteken és a limfoblasztokon keresztül vezet az útjuk (lásd a Sejtes védekezés 1. ábráját). A T-sejtekkel ellentétben azonban a B-sejtekké válni hivatott limfoblasztok nem hagyják el a csontvelőt, és nem utaznak a tímuszba érés céljából. Az esetleges B-sejtek inkább a csontvelőben érnek tovább.
A B-sejtek érésének első lépése az antigénkötő receptoraik működőképességének felmérése. Ez a normális funkcionális receptorokkal rendelkező B-sejtek pozitív szelekciója révén történik. Ezt követően a negatív szelekció mechanizmusa az önreagáló B-sejtek eliminálására és az autoimmunitás kockázatának minimalizálására szolgál. Az önreagáló B-sejtek negatív szelekciója magában foglalhatja az apoptózis általi eliminációt, a receptorok szerkesztését vagy módosítását, hogy azok többé ne legyenek önreaktívak, vagy anergia indukálását a B-sejtben. A csontvelőben a szelekción átesett éretlen B-sejtek ezután a lépbe kerülnek az érés végső szakaszába. Ott válnak naiv érett B-sejtekké, azaz, érett B-sejtek, amelyek még nem aktiválódtak.
Gondolj bele
- Hasonlítsuk össze a B-sejtek érését a T-sejtek érésével.
B-sejt receptorok
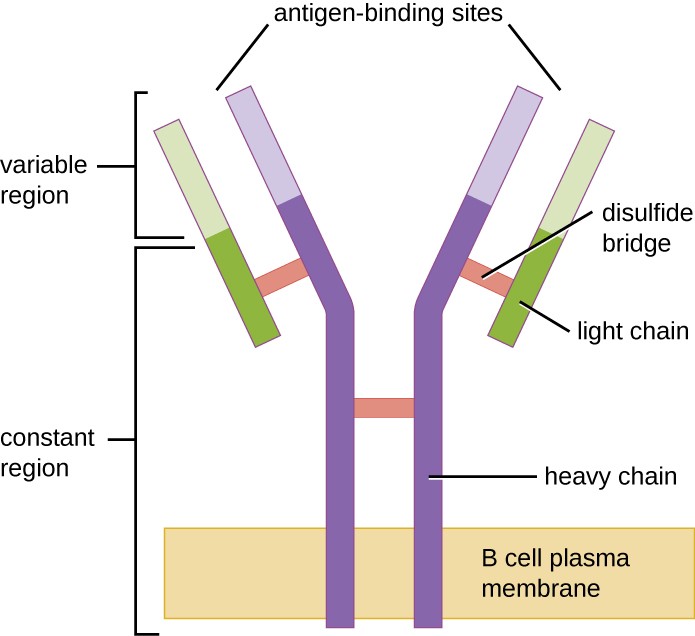
1. ábra. A B-sejt receptorok a B-sejtek membránjába ágyazódnak. Az egyetlen sejtben lévő összes receptor változó régiója ugyanazt a specifikus antigént köti.
A T-sejtekhez hasonlóan a B-sejtek is rendelkeznek különböző specifitású antigénspecifikus receptorokkal. Bár optimális működésükhöz a T-sejtekre támaszkodnak, a B-sejtek a T-sejtek segítsége nélkül is aktiválhatók. A naiv érett B-sejtek B-sejt receptorai (BCR) az IgD és IgM membránhoz kötött monomer formái. Két azonos nehézláncból és két azonos könnyű láncból állnak, amelyeket diszulfidkötések kötnek össze “Y” alapformába (1. ábra). Az Y alakú molekula törzse, a két nehézlánc állandó régiója a B-sejtmembránon átível. A B-sejt külseje felé kitett két antigénkötő hely a specifikus patogén epitópok megkötésében vesz részt az aktivációs folyamat elindítása érdekében. Becslések szerint minden naiv, érett B-sejt membránján több mint 100 000 BCR található, és ezek mindegyike azonos epitópkötő specifitással rendelkezik.
Azért, hogy felkészüljenek a mikrobiális epitópok széles skálájára való reagálásra, a B-sejtek a T-sejtekhez hasonlóan több száz génszakasz genetikai átrendeződésével biztosítják a receptorspecifikumok szükséges változatosságát. A BCR nehézláncának változó régiója a TCR β-láncához hasonlóan V, D és J szegmensekből áll. A BCR könnyű láncának változó régiója V és J szegmensekből áll, hasonlóan a TCR α láncához. A V-J-D (nehézlánc) és V-J (könnyűlánc) minden lehetséges kombinációjának genetikai átrendeződése milliónyi egyedi antigénkötő helyet biztosít a BCR és az aktiválódás után szekretált antitestek számára.
A BCR-ek és a TCR-ek közötti egyik fontos különbség az, ahogyan kölcsönhatásba léphetnek az antigén epitópokkal. Míg a TCR-ek csak olyan antigén epitópokkal tudnak kölcsönhatásba lépni, amelyek az MHC I vagy MHC II antigénkötő hasadékában vannak jelen, addig a BCR-eknek nincs szükségük az MHC-vel történő antigénprezentációra; kölcsönhatásba léphetnek a szabad antigéneken lévő epitópokkal vagy az intakt kórokozók felszínén megjelenő epitópokkal. Egy másik fontos különbség, hogy a TCR-ek csak fehérjeepitópokat ismernek fel, míg a BCR-ek különböző molekuláris osztályokhoz (pl. fehérjékhez, poliszacharidokhoz, lipopoliszacharidokhoz) kapcsolódó epitópokat ismerhetnek fel.
A B-sejtek aktiválása az antigén molekuláris osztályától függően különböző mechanizmusokon keresztül történik. A B-sejt fehérjeantigén általi aktiválása megköveteli, hogy a B-sejt APC-ként működjön, és a fehérjeepitópokat MHC II-vel mutassa be a segítő T-sejteknek. Mivel a B-sejtek aktiválása a T-sejtektől függ, a fehérjeantigéneket a T-függő antigének közé sorolják. Ezzel szemben a poliszacharidokat, lipopoliszacharidokat és más nem fehérje antigéneket T-független antigéneknek tekintik, mivel antigénfeldolgozás és T-sejteknek történő prezentáció nélkül is képesek aktiválni a B-sejteket.
Gondolj bele
- Milyen típusú molekulák szolgálnak BCR-ként?
- Mi a különbség a TCR-ek és a BCR-ek között az antigénfelismerés tekintetében?
- Mely molekulaosztályok T-függő antigének és melyek T-független antigének?
A B-sejtek T-sejt-független aktivációja
A B-sejteknek a segítő T-sejtek együttműködése nélküli aktivációját T-sejt-független aktivációnak nevezzük, és akkor következik be, amikor a BCR-ek T-független antigénekkel lépnek kölcsönhatásba. A T-független antigének (pl. poliszacharid kapszulák, lipopoliszacharid) szerkezetében ismétlődő epitóp egységek vannak, és ez az ismétlődés lehetővé teszi több BCR keresztkötését, ami az aktiváció első jelét adja (2. ábra). Mivel a T-sejtek nem vesznek részt, a második jelnek más forrásból kell származnia, például a toll-szerű receptorok kölcsönhatásaiból a PAMP-okkal vagy a komplementrendszer faktorainak kölcsönhatásaiból.
Amikor egy B-sejt aktiválódik, klonális proliferáción megy keresztül, és a leánysejtek plazmasejtekké differenciálódnak. A plazmasejtek antitestgyárak, amelyek nagy mennyiségű antitestet választanak ki. A differenciálódás után a felszíni BCR-ek eltűnnek, és a plazmasejt pentamer IgM molekulákat választ ki, amelyeknek ugyanaz az antigénspecificitása, mint a BCR-eknek (2. ábra).
A T-sejt-független válasz rövid életű, és nem eredményezi memória B-sejtek termelődését. Így nem eredményez másodlagos választ a T-független antigénekkel való későbbi expozíciókra.

2. ábra. A T-független antigének olyan ismétlődő epitópokkal rendelkeznek, amelyek a T-sejtek közreműködése nélkül is képesek B-sejt felismerést és aktivációt kiváltani. A B-sejt aktiválásához egy második jel, például a TLR-ek és a PAMP-ok kölcsönhatása (nem látható) is szükséges. Miután aktiválódott, a B-sejt proliferálódik és antitest-szekretáló plazmasejtekké differenciálódik.
Gondolj bele
- Mi az a két jel, amely a B-sejtek T-sejt-független aktiválásához szükséges?
- Mi a plazmasejt funkciója?
A B-sejtek T-sejt-függő aktivációja

3. ábra. Kattintson a nagyobb képért. A B-sejtek T-sejt-függő aktivációja során a B-sejt felismer és internalizál egy antigént, majd bemutatja azt egy segítő T-sejtnek, amely ugyanarra az antigénre specifikus. A segítő T-sejt kölcsönhatásba lép a B-sejt által prezentált antigénnel, ami aktiválja a T-sejtet és serkenti a citokinek felszabadulását, amelyek aztán aktiválják a B-sejtet. A B-sejt aktivációja kiváltja a B-sejtek proliferációját és differenciálódását B-sejtekké és plazmasejtekké.
A B-sejtek T-sejt-függő aktivációja összetettebb, mint a T-sejt-független aktiváció, de a kialakuló immunválasz erősebb és memóriát fejleszt. A T-sejt-függő aktiváció történhet szabad fehérjeantigénekre vagy ép kórokozóhoz kapcsolódó fehérjeantigénekre adott válaszként. A naiv érett B-sejt BCR-jei és a szabad fehérjeantigén közötti kölcsönhatás serkenti az antigén internalizációját, míg az intakt kórokozóval társult antigénekkel való kölcsönhatás az antigénnek a kórokozóból való kivonását indítja el az internalizáció előtt. Miután internalizálódott a B-sejt belsejében, a fehérjeantigént feldolgozzák és az MHC II-vel prezentálják. A prezentált antigént ezután az ugyanarra az antigénre specifikus segítő T-sejtek felismerik. A segítő T-sejt TCR-je felismeri az idegen antigént, és a T-sejt CD4 molekulája kölcsönhatásba lép a B-sejt MHC II-vel. Az ugyanarra az antigénre specifikus B-sejtek és segítő T-sejtek közötti koordinációt nevezzük kapcsolt felismerésnek.
Amint a kapcsolt felismerés aktiválja őket, a TH2 sejtek olyan citokineket termelnek és választanak ki, amelyek aktiválják a B-sejtet és klonális leánysejtekké való proliferációt okoznak. A proliferáció több körét követően a TH2 sejtek által biztosított további citokinek serkentik az aktivált B-sejtklónok differenciálódását memória B-sejtekké, amelyek gyorsan reagálnak az ugyanazon fehérjeepitópnak való későbbi expozíciókra, valamint plazmasejtekké, amelyek elveszítik membrán BCR-jeiket és kezdetben pentamer IgM-et választanak ki (3. ábra).
Az IgM kezdeti szekrécióját követően a TH2 sejtek által szekretált citokinek stimulálják a plazmasejteket, hogy az IgM termelésről IgG, IgA vagy IgE termelésre váltsanak. Ez az osztályváltásnak vagy izotípusváltásnak nevezett folyamat lehetővé teszi, hogy az ugyanabból az aktivált B-sejtből klónozott plazmasejtek különböző, azonos epitópspecifitású antitestosztályokat termeljenek. Az osztályváltás az antitest osztályát meghatározó konstans régiót kódoló génszakaszok genetikai átrendeződésével valósul meg. A változó régió nem változik, így az új antitestosztály megtartja az eredeti epitópspecificitást.
Gondolj bele
- Milyen lépések szükségesek a B-sejtek T-sejt-függő aktiválásához?
- Mi az antitest osztályváltás és miért fontos?
Primer és másodlagos válaszok
A B-sejtek T-sejt-függő aktivációja fontos szerepet játszik mind az adaptív immunitással kapcsolatos primer, mind a másodlagos válaszokban. A fehérjeantigénnel való első érintkezéskor T-sejt-függő primer antitestválasz lép fel. Az elsődleges válasz kezdeti szakasza egy kb. 10 napos késleltetési vagy látens időszak, amely alatt a szérumban nem mutatható ki antitest. Ez a késleltetési időszak az elsődleges válasz összes lépéséhez szükséges idő, beleértve az antigén BCR-ekkel való naiv, érett B-sejt kötődését, az antigén feldolgozását és prezentációját, a segítő T-sejtek aktiválódását, a B-sejtek aktiválódását és a klonális proliferációt. A késleltetési időszak végét a szérum IgM-szintjének emelkedése jellemzi, mivel a TH2 sejtek stimulálják a B-sejtek plazmasejtekké történő differenciálódását. Az IgM-szintek az elsődleges antigén-expozíciót követően körülbelül 14 nappal érik el csúcspontjukat; körülbelül ugyanebben az időben a TH2 stimulálja az antitestosztály-váltást, és a szérum IgM-szintje csökkenni kezd. Eközben az IgG szintje növekszik, amíg az elsődleges válasz után körülbelül három héttel el nem éri a csúcsot (4. ábra).
Az elsődleges válasz során a klónozott B-sejtek egy része memória-B-sejtekké differenciálódik, amelyeket úgy programoztak, hogy a későbbi expozíciókra reagáljanak. Ez a másodlagos válasz gyorsabban és erőteljesebben jelentkezik, mint az elsődleges válasz. A késleltetési idő mindössze néhány napra csökken, és az IgG-termelés jelentősen magasabb, mint az elsődleges válasz esetében megfigyelt (4. ábra). Ezenkívül a másodlagos válasz során termelődő antitestek hatékonyabbak és nagyobb affinitással kötődnek a célzott epitópokhoz. A másodlagos válasz során termelődő plazmasejtek tovább élnek, mint az elsődleges válasz során termelődők, így a specifikus antitestek szintje hosszabb ideig emelkedett marad.

4. ábra. Az elsődleges válaszhoz képest a másodlagos antitestválasz gyorsabban jelentkezik, és magasabb és tartósabb antitestszintet produkál. A másodlagos válasz többnyire IgG-t érint.
Gondolj bele
- Milyen események történnek az elsődleges antitestválasz késleltetési időszakában?
- Miért marad az ellenanyagszint hosszabb ideig emelkedett a másodlagos ellenanyagválasz során?
Főbb fogalmak és összefoglalás
- A B-limfociták vagy B-sejtek termelik a humorális immunitásban részt vevő ellenanyagokat. A B-sejtek a csontvelőben termelődnek, ahol az érés kezdeti szakaszai történnek, és a lépbe vándorolnak az érés utolsó lépéseihez, ahol naiv, érett B-sejtekké válnak.
- A B-sejt receptorok (BCR) az IgD és IgM membránhoz kötött monomer formái, amelyek Fab antigénkötő régióikkal specifikus antigén epitópokat kötnek. Az antigénkötési specifitás diverzitása a V, D és J szegmensek genetikai átrendeződésével jön létre, hasonlóan a TCR diverzitáshoz használt mechanizmushoz.
- A fehérjeantigéneket T-függő antigéneknek nevezik, mert csak a segítő T-sejtek együttműködésével képesek aktiválni a B-sejteket. Más molekulaosztályok nem igényelnek T-sejt együttműködést, és azokat T-független antigéneknek nevezzük.
- A B-sejtek T-sejt-független aktiválása a BCR-ek keresztkötését jelenti a repetitív nem fehérje antigén epitópok által. Jellemzője az IgM plazmasejtek általi termelése, és nem hoz létre memória B-sejteket.
- A B-sejtek T-sejt-függő aktivációja magában foglalja a fehérje antigének feldolgozását és bemutatását a segítő T-sejtek számára, a B-sejtek aktiválását az aktivált TH2 sejtek által szekretált citokinek által, és a plazmasejteket, amelyek az osztályváltás eredményeként különböző osztályú antitesteket termelnek. Memória B-sejtek is termelődnek.
- A T-függő antigéneknek való másodlagos kitettség a memória B-sejtek által indított másodlagos antitestválaszt eredményez. A másodlagos válasz gyorsabban fejlődik ki, és magasabb és tartósabb antitestszintet termel, nagyobb affinitással a specifikus antigénhez.
Multiple Choice
Az alábbiak közül melyik lenne T-függő antigén?
- lipopolysaccharide
- glycolipid
- protein
- carbohydrate
Which of the following would be a BCR?
- CD4
- MHC II
- MHC I
- IgD
Which of the following does not occur during the lag period of the primary antibody response?
- activation of helper T cells
- class switching to IgG
- presentation of antigen with MHC II
- binding of antigen to BCRs
Fill in the Blank
________ antigens can stimulate B cells to become activated but require cytokine assistance delivered by helper T cells.
T-independent antigens can stimulate B cells to become activated and secrete antibodies without assistance from helper T cells. These antigens possess ________ antigenic epitopes that cross-link BCRs.
Think about It
A patient lacks the ability to make functioning T cells because of a genetic disorder. Would this patient’s B cells be able to produce antibodies in response to an infection? Explain your answer.