いったん相互リン酸化されると、RTK の細胞質尾部は、シグナル伝達に関わるさまざまな細胞内タンパク質のドッキング プラットフォームとして機能します。 これらのタンパク質は、細胞質RTK受容体尾部のリン酸化チロシンに結合する、SH2と呼ばれる特殊なドメインを持っている。 活性化されたRTKには、複数のSH2含有タンパク質が同時に結合することができ、複数の細胞内シグナル伝達経路を同時に活性化することが可能である。 最終的には、RTKの活性化により、遺伝子の転写に変化が生じる。 シグナルが膜から核に移動する際、細胞内の様々なシグナル伝達経路の中間体間のクロストークにより、シグナル伝達が複雑化する(図1)。

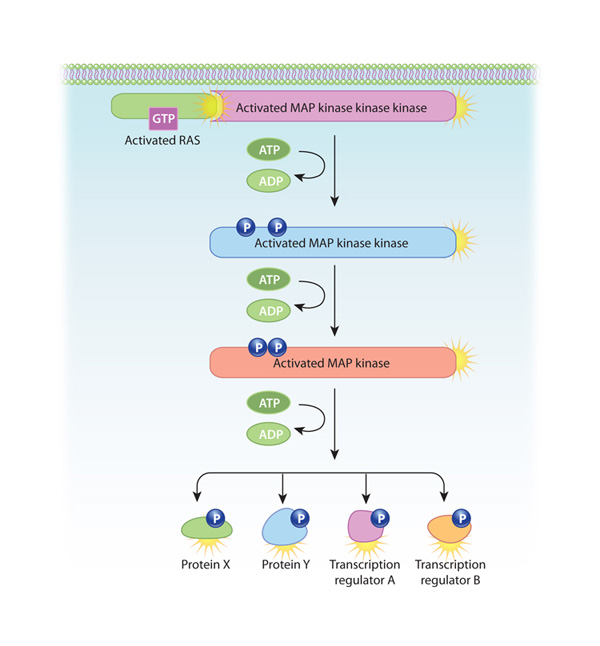
One of the most common intracellular signaling pathways triggered by RTKs is known as the mitogen-activated protein (MAP) kinase cascade, because it involves three serine-threonine kinases. この経路は、細胞膜に固定された小さなGタンパク質であるRasが活性化されることから始まる。 Rasは細胞膜に固定された小さなGタンパク質であり、不活性な状態ではGDPと結合している。 しかし、SH2含有タンパク質が活性化したRTKに結合すると、RasはGDPの代わりにGTPを結合し、活性化される。 次に、GTP結合したRas(それ自身はキナーゼではない)は、MAPキナーゼカスケードの最初のセリン・スレオニンキナーゼを活性化する。 このカスケード内の3つのキナーゼはそれぞれ、次のキナーゼをリン酸化することにより活性化する。 この経路の3つのキナーゼはすべて複数の基質をリン酸化するため、最初のシグナルは各ステップで増幅される。 そして、この経路の最後の酵素が転写調節因子をリン酸化することで、遺伝子の転写が変化する(図2)。 神経成長因子や血小板由来成長因子など、多くの成長因子がこの経路を利用している。
すべてのRTKがMAPキナーゼのカスケードを用いて核に情報を送るわけではない。 例えば、インスリン様成長因子受容体はホスホイノシチド3キナーゼという酵素を活性化し、細胞膜のイノシトールリン脂質をリン酸化して、(MAPキナーゼカスケードとは異なる)タンパク質キナーゼカスケードにつながり、核にシグナルが伝達されるのである。 他のRTKは、より直接的な経路で核に到達する。 STATタンパク質として知られる転写調節因子(signal transducers and activators of transcriptionの頭文字をとったもの)は、サイトカインやいくつかのホルモンの受容体にあるリン酸化チロシンに結合する。 STATタンパク質は、活性化されると核内に直接移動し、転写に変化をもたらす。