Lärandemål
- Beskriv produktion och mognad av B-celler
- Genomföra strukturen av B-cellsreceptorer och T-cellsreceptorer
- Genomföra T-beroende och T-oberoende aktivering av B-celler
- Jämför primära och sekundära antikroppssvar
Humoral immunitet hänvisar till mekanismer i det adaptiva immunförsvaret som förmedlas av antikroppar som utsöndras av B-lymfocyter, eller B-celler. Detta avsnitt kommer att fokusera på B-celler och diskutera deras produktion och mognad, receptorer och aktiveringsmekanismer.
B-cellers produktion och mognad
Likt T-celler bildas B-celler från multipotenta hematopoietiska stamceller (HSC) i benmärgen och följer en väg via lymfoid stamcell och lymfoblast (se figur 1 i Cellular Defenses). Till skillnad från T-celler lämnar dock inte lymfoblaster som är avsedda att bli B-celler benmärgen och reser till thymus för mognad. Eventuella B-celler fortsätter snarare att mogna i benmärgen.
Det första steget i B-cellernas mognad är en bedömning av funktionaliteten hos deras antigenbindande receptorer. Detta sker genom positivt urval av B-celler med normalt fungerande receptorer. En mekanism för negativt urval används sedan för att eliminera självreagerande B-celler och minimera risken för autoimmunitet. Negativt urval av självreagerande B-celler kan innebära eliminering genom apoptos, redigering eller modifiering av receptorerna så att de inte längre är självreaktiva, eller framkallande av anergi i B-cellen. Omogna B-celler som klarar urvalet i benmärgen reser sedan till mjälten för sina sista mognadsstadier. Där blir de naiva mogna B-celler, dvs, mogna B-celler som ännu inte har aktiverats.
Tänk efter
- Genomföra mognaden av B-celler med mognaden av T-celler.
B-cellreceptorer
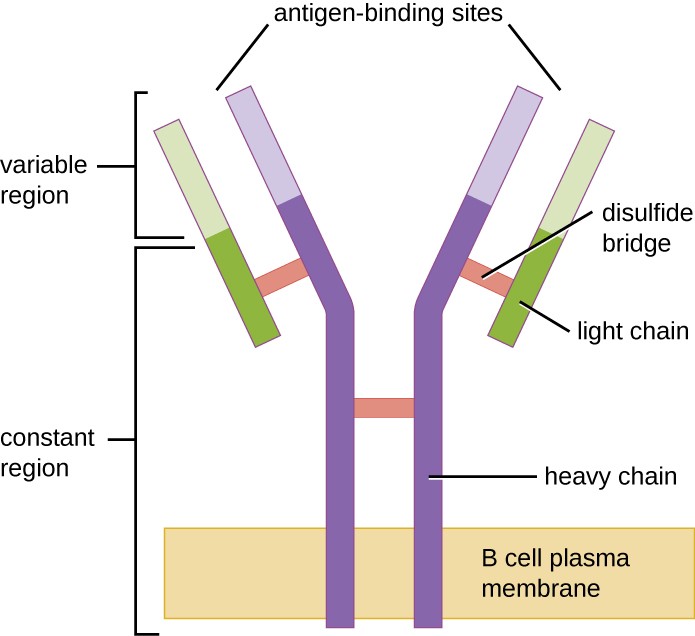
Figur 1. B-cellsreceptorer är inbäddade i B-cellernas membran. De variabla regionerna hos alla receptorer på en enskild cell binder samma specifika antigen.
Likt T-celler har B-celler antigenspecifika receptorer med olika specificiteter. Även om de är beroende av T-celler för att fungera optimalt kan B-celler aktiveras utan hjälp från T-celler. B-cellsreceptorer (BCR) för naiva mogna B-celler är membranbundna monomera former av IgD och IgM. De har två identiska tunga kedjor och två identiska lätta kedjor som är förbundna med disulfidbindningar i en grundläggande Y-form (figur 1). Den Y-formade molekylens stam, den konstanta regionen av de två tunga kedjorna, sträcker sig över B-cellmembranet. De två antigenbindningsställen som är exponerade på B-cellens utsida är involverade i bindningen av specifika patogena epitoper för att inleda aktiveringsprocessen. Man uppskattar att varje naiv, mogen B-cell har uppemot 100 000 BCR på sitt membran, och var och en av dessa BCR har en identisk epitopbindande specificitet.
För att vara beredd att reagera på ett brett spektrum av mikrobiella epitoper använder sig B-celler, liksom T-celler, av en genetisk omgruppering av hundratals genavsnitt för att åstadkomma den nödvändiga mångfalden av receptorspecificiteter. Den variabla regionen i BCR:s tunga kedja består av V-, D- och J-segmenten, vilket liknar TCR:s β-kedja. Den variabla regionen i den lätta BCR-kedjan består av V- och J-segmenten, som liknar TCR:s α-kedja. Genetisk omarrangemang av alla möjliga kombinationer av V-J-D (tung kedja) och V-J (lätt kedja) ger miljontals unika antigenbindningsställen för BCR och för de antikroppar som utsöndras efter aktivering.
En viktig skillnad mellan BCR och TCR är hur de kan interagera med antigena epitoper. Medan TCR:er endast kan interagera med antigena epitoper som presenteras inom den antigenbindande klyftan i MHC I eller MHC II, kräver BCR:er inte antigenpresentation med MHC; de kan interagera med epitoper på fria antigener eller med epitoper som visas på ytan av intakta patogener. En annan viktig skillnad är att TCR:er endast känner igen proteinepitoper, medan BCR:er kan känna igen epitoper som är associerade med olika molekylära klasser (t.ex. proteiner, polysackarider, lipopolysackarider).
Aktivering av B-celler sker genom olika mekanismer beroende på antigenets molekylära klass. Aktivering av en B-cell av ett proteinantigen kräver att B-cellen fungerar som en APC och presenterar proteinepitopen med MHC II för hjälpande T-celler. På grund av deras beroende av T-celler för aktivering av B-celler klassificeras proteinantigener som T-beroende antigener. Polysackarider, lipopolysackarider och andra icke-proteinantigener betraktas däremot som T-oberoende antigener eftersom de kan aktivera B-celler utan antigenbearbetning och presentation till T-celler.
Tänk efter
- Vilka typer av molekyler tjänar som BCR?
- Vilka skillnader finns det mellan TCR:er och BCR:er med avseende på antigenigenigenkännedom?
- Vilka molekylklasser är T-beroende antigener och vilka är T-oberoende antigener?
T-celloberoende aktivering av B-celler
Aktivering av B-celler utan samarbete med hjälpande T-celler kallas T-celloberoende aktivering och inträffar när BCR:er interagerar med T-oberoende antigener. T-oberoende antigener (t.ex. polysackaridkapslar, lipopolysackarid) har repetitiva epitopenheter i sin struktur, och denna repetition möjliggör tvärbindning av flera BCR, vilket ger den första signalen för aktivering (figur 2). Eftersom T-celler inte är inblandade måste den andra signalen komma från andra källor, t.ex. interaktioner mellan toll-liknande receptorer och PAMPs eller interaktioner med faktorer från komplementsystemet.
När en B-cell väl är aktiverad genomgår den en klonal proliferation och dottercellerna differentieras till plasmaceller. Plasmaceller är antikroppsfabriker som utsöndrar stora mängder antikroppar. Efter differentieringen försvinner BCR:erna på ytan och plasmacellen utsöndrar pentameriska IgM-molekyler som har samma antigenspecificitet som BCR:erna (figur 2).
Det T-celloberoende svaret är kortvarigt och resulterar inte i produktion av minnes-B-celler. Det kommer därför inte att resultera i ett sekundärt svar vid efterföljande exponering för T-oberoende antigener.

Figur 2. T-oberoende antigener har repetitiva epitoper som kan framkalla B-cellsigenkänning och aktivering utan inblandning av T-celler. En andra signal, t.ex. interaktion mellan TLRs och PAMPs (ej visad), krävs också för aktivering av B-cellen. När den väl är aktiverad prolifererar B-cellen och differentieras till antikroppsutsöndrande plasmaceller.
Tänk efter
- Vad är de två signaler som krävs för T-cellsoberoende aktivering av B-celler?
- Vad är en plasmacells funktion?
T-cellberoende aktivering av B-celler

Figur 3. Klicka för en större bild. Vid T-cellsberoende aktivering av B-celler känner B-cellen igen och internaliserar ett antigen och presenterar det för en hjälpande T-cell som är specifik för samma antigen. Den hjälpsamma T-cellen interagerar med antigenet som presenteras av B-cellen, vilket aktiverar T-cellen och stimulerar frisättningen av cytokiner som sedan aktiverar B-cellen. Aktiveringen av B-cellen utlöser proliferation och differentiering till B-celler och plasmaceller.
T-cellberoende aktivering av B-celler är mer komplex än T-celloberoende aktivering, men det resulterande immunsvaret är starkare och utvecklar minne. T-cellsberoende aktivering kan ske antingen som svar på fria proteinantigener eller på proteinantigener som är associerade med en intakt patogen. Interaktion mellan BCR:erna på en naiv mogen B-cell och ett fritt proteinantigen stimulerar internalisering av antigenet, medan interaktion med antigener som är associerade med en intakt patogen initierar extraktionen av antigenet från patogenet före internalisering. När det har internaliserats i B-cellen bearbetas proteinantigenet och presenteras med MHC II. Det presenterade antigenet känns sedan igen av hjälpande T-celler som är specifika för samma antigen. Den hjälpsamma T-cellens TCR känner igen det främmande antigenet och T-cellens CD4-molekyl interagerar med MHC II på B-cellen. Samordningen mellan B-celler och hjälpande T-celler som är specifika för samma antigen kallas länkat erkännande.
När de aktiveras av länkat erkännande producerar och utsöndrar TH2-celler cytokiner som aktiverar B-cellen och orsakar proliferation till klonala dotterceller. Efter flera omgångar av proliferation stimulerar ytterligare cytokiner från TH2-cellerna differentieringen av aktiverade B-cellskloner till minnes-B-celler, som snabbt reagerar på efterföljande exponering för samma proteinepitop, och plasmaceller som förlorar sina BCR-membran och initialt utsöndrar pentameriskt IgM (figur 3).
Efter den initiala utsöndringen av IgM stimulerar cytokiner som utsöndras av TH2-celler plasmacellerna att övergå från IgM-produktion till produktion av IgG, IgA eller IgE. Denna process, som kallas klassväxling eller isotypväxling, gör att plasmaceller som klonats från samma aktiverade B-cell kan producera en mängd olika antikroppsklasser med samma epitopspecificitet. Klassväxling sker genom genetisk omläggning av gensegment som kodar för den konstanta regionen, som bestämmer en antikroppsklass. Den variabla regionen ändras inte, så den nya klassen av antikroppar behåller den ursprungliga epitopspecificiteten.
Tänk efter
- Vilka steg krävs för T-cellsberoende aktivering av B-celler?
- Vad är byte av antikroppsklass och varför är det viktigt?
Primär och sekundär respons
T-cellberoende aktivering av B-celler spelar en viktig roll i både den primära och sekundära responsen i samband med adaptiv immunitet. Vid den första exponeringen för ett proteinantigen uppstår ett T-cellsberoende primärt antikroppssvar. Det första skedet av det primära svaret är en fördröjningsperiod, eller latent period, på cirka 10 dagar, under vilken inga antikroppar kan påvisas i serum. Denna eftersläpningsperiod är den tid som krävs för alla steg i det primära svaret, inklusive naiva mogna B-cellers bindning av antigen med BCR, antigenbearbetning och antigenpresentation, aktivering av T-hjälparceller, aktivering av B-celler och klonproliferation. Slutet av lagringstiden kännetecknas av en ökning av IgM-nivåerna i serum, eftersom TH2-cellerna stimulerar B-cellernas differentiering till plasmaceller. IgM-nivåerna når sin topp cirka 14 dagar efter den primära antigenexponeringen. Ungefär vid samma tidpunkt stimulerar TH2- cellerna byte av antikroppsklass och IgM-nivåerna i serum börjar sjunka. Samtidigt ökar IgG-nivåerna tills de når en topp ungefär tre veckor efter det primära svaret (figur 4).
Under det primära svaret differentieras en del av de klonade B-cellerna till minnes-B-celler som är programmerade att svara på efterföljande exponeringar. Detta sekundära svar sker snabbare och kraftigare än det primära svaret. Fördröjningsperioden minskar till endast några dagar och produktionen av IgG är betydligt högre än vad som observerades för det primära svaret (figur 4). Dessutom är de antikroppar som produceras under det sekundära svaret effektivare och binder med högre affinitet till de målinriktade epitopen. Plasmaceller som produceras under sekundära svar lever längre än de som produceras under det primära svaret, så nivåerna av specifika antikroppar förblir förhöjda under en längre tid.

Figur 4. Jämfört med det primära svaret uppträder det sekundära antikroppssvaret snabbare och ger antikroppsnivåer som är högre och mer långvariga. Det sekundära svaret involverar främst IgG.
Tänk efter
- Vilka händelser inträffar under fördröjningsperioden för det primära antikroppssvaret?
- Varför förblir antikroppsnivåerna förhöjda längre under det sekundära antikroppssvaret?
Nyckelbegrepp och sammanfattning
- B-lymfocyter eller B-celler producerar antikroppar som är involverade i humoral immunitet. B-celler produceras i benmärgen, där de första stadierna av mognad sker, och reser till mjälten för de sista stadierna av mognad till naiva mogna B-celler.
- B-cellreceptorer (BCR) är membranbundna monomera former av IgD och IgM som binder specifika antigenepitoper med sina Fab-antigenbindande regioner. Mångfald av antigenbindningsspecificitet skapas genom genetiska omarrangemang av V-, D- och J-segmenten på samma sätt som den mekanism som används för TCR-mångfald.
- Proteinantigen kallas T-beroende antigener eftersom de endast kan aktivera B-celler med hjälp av hjälpartade T-celler. Andra molekylklasser kräver inte samarbete med T-cellerna och kallas T-oberoende antigener.
- T-celloberoende aktivering av B-celler innebär korsbindning av BCR:er med repetitiva icke-proteinantigenepitoper. Den kännetecknas av produktion av IgM av plasmaceller och ger inte upphov till minnes-B-celler.
- T-cellberoende aktivering av B-celler inbegriper bearbetning och presentation av proteinantigener till hjälpartade T-celler, aktivering av B-cellerna genom cytokiner som utsöndras från aktiverade TH2-celler och plasmaceller som producerar olika klasser av antikroppar som ett resultat av klassväxling. Minnes-B-celler produceras också.
- Sekundär exponering för T-beroende antigener resulterar i ett sekundärt antikroppssvar som initieras av minnes-B-celler. Det sekundära svaret utvecklas snabbare och producerar högre och mer ihållande nivåer av antikroppar med högre affinitet för det specifika antigenet.
Multiple Choice
Vilket av följande skulle vara ett T-beroende antigen?
- lipopolysaccharide
- glycolipid
- protein
- carbohydrate
Which of the following would be a BCR?
- CD4
- MHC II
- MHC I
- IgD
Which of the following does not occur during the lag period of the primary antibody response?
- activation of helper T cells
- class switching to IgG
- presentation of antigen with MHC II
- binding of antigen to BCRs
Fill in the Blank
________ antigens can stimulate B cells to become activated but require cytokine assistance delivered by helper T cells.
T-independent antigens can stimulate B cells to become activated and secrete antibodies without assistance from helper T cells. These antigens possess ________ antigenic epitopes that cross-link BCRs.
Think about It
A patient lacks the ability to make functioning T cells because of a genetic disorder. Would this patient’s B cells be able to produce antibodies in response to an infection? Explain your answer.