1. Översikt över Erk-signalering
Den extracellulära signalreglerade kinasen (ERK) är en av de viktigaste signalkassetterna i den mitogenaktiverade proteinkinasets (MAPK) signalväg. ERK-kaskaden aktiveras av en mängd olika extracellulära ämnen, inklusive tillväxtfaktorer, hormoner och även cellulära påfrestningar för att inducera cellulära processer som huvudsakligen omfattar proliferation och differentiering, men under vissa förhållanden även stressrespons och andra. De viktigaste komponenterna i MAPK/ERK-kinaset (MEKK) är Raf-familjemedlemmarna Raf-1, A-Raf och B-Raf som fosforylerar två serinrester på MAPK/ERK-kinaset (MEK)-komponenterna MEK1/2. ERK1/2 är ansvariga för att stimulera nedströms effektorer, varav många är transkriptionsfaktorer. En av de viktigaste transkriptionsfaktorerna är cyclic AMP response element-binding protein (CREB).
Denna ERK-bana bidrar till kontrollen av ett stort antal cellulära processer:
– Reglering av cellproliferation, t.ex. aktivering av T-celler
– Synaptisk plasticitet, t.ex. lång-långtidspotentiering (LTP) i hippocampala neuroner
– Proliferation av endotelceller under angiogenes
– Fosforylering av transkriptionsfaktorn p53
– Omformning av ERK-signalvägen kan bidra till utvecklingen av polycystisk njursjukdom
– Aktivering av fosfolipas A2 (PLA2) i mastceller.
2. Erk-signalkaskad
Aktiveringen av ERK1/2-kaskaden initieras oftast vid membranreceptorer, såsom receptor-Tyr-kinaser (RTK), G-protein-kopplade receptorer (GPCR), jonkanaler och andra. Dessa receptorer överför signalen genom att rekrytera adaptorproteiner (t.ex. Grb2) och utbytesfaktorer (t.ex. SOS) som i sin tur inducerar aktivering av Ras vid plasmamembranen eller membranen i andra organeller. Det aktiverade, GTP-bundna Ras överför sedan signalen genom att aktivera proteinkinaserna Raf-1, B-Raf och A-Raf (Rafs) inom MAP3K-nivån i denna kaskad. Denna aktivering sker genom att Rafs rekryteras till membranen, där de sedan fosforyleras och aktiveras. MOS är en annan MAP3K i ERK-kaskaden, men den verkar huvudsakligen i fortplantningssystemet genom ett distinkt regleringssätt. Från MAP3K-nivån överförs signalen sedan nedåt i kaskaden genom MAPKK-komponenterna: MEK1/2. De aktiveras genom serinfosforylering vid det MAPKK-typiska Ser-Xaa-Ala-Xaa-Ser/Thr-motivet i deras aktiveringsloop. MEK1/2 aktiverar i sin tur sina enda kända substrat, naturliga ERK1/2, som fungerar som deras enda nedströmsmål, vilket tyder på att MEK1/2 fungerar som de specificitetsbestämmande komponenterna i ERK1/2-kaskaden. MEK1/2 är de enda dubbelspecifika proteinkinaser som kan fosforylera både reglerande Thr- och Tyr-rester av ERK1/2. Omkring 200 olika substrat av ERK1/2 har hittills identifierats. De är substrat antingen i cytosolen (t.ex. PLA2 RSK) eller vid translokation i kärnan, inklusive transkriptionsfaktorer som Elk1, c-Fos och c-Jun.
– MAP3K-Raf 1
Raf-1 är den bäst studerade av tillväxtfaktorsignalmolekylerna på MAP3K-nivån i kaskaden. Det är ett 70-75 kDa protein serin/treoninkinas som innehåller en kinasdomän i den COOH-terminala halvan av molekylen och en regulatorisk domän som upptar resten av molekylen. Vid stimulering med olika mitogener genomgår Raf-1 en övergående aktivering inom 2-3 mm. Mekanismen för aktivering av Raf-1 har studerats ingående under de senaste åren. Det står nu klart att Raf-1 ligger nedströms från HAS, som verkar interagera direkt med den NH2-terminala delen av Raf-1 vid stimulering.
– MAPKK-MEK1/2
MEK:erna utgör en evolutionärt bevarad familj av proteinserin/treoninkinaser som hittills omfattar tre mycket homologa (85 %) däggdjursisoformer. Mekanismen för MEK-1-aktivering inbegriper proteinfosforylering endast på serinrester och verkar vara likartad oavsett om den fosforyleras av MEKK, Raf-1 eller genom autofosforylering, vilket fastställs med hjälp av foskopeptidkartor. MAPKK (MEK) är mycket specifika för nedströms-komponenterna ERK-1 och ERK-2. Dessutom kunde MAPKK inte känna igen vare sig den denaturerade formen av sitt substrat eller peptider som innehåller fosforyleringsställena i ERK, vilket tyder på att enzymet kräver MAPK:s nativa form.
– MAPK-ERK-1/2
På grund av sin höga grad av likhet anses ERK-1 och ERK-2 vanligen vara funktionellt redundanta. Den viktigaste uppströmsmekanismen som leder till fosforylering av ERK-1 och -2 på båda dessa reglerande rester är deras fosforylering av MEK. På grund av den ganska breda karaktären hos dess substratigenkänning kan ERK fosforylera ett stort antal proteiner efter deras aktivering. Dessa proteiner är ofta av reglerande karaktär och finns både i cytoplasman och i kärnan.
3. Nedströms signalering av Erk-banan
– Cellproliferation
Aktiverad ERK1/2 fosforylerar RSK och både RSK och ERK translokaliseras till kärnan där de aktiverar flera transkriptionsfaktorer som CREB, Fos och Elk-1, vilket i slutändan resulterar i syntesen av effectorproteiner och orsakar förändringar i cellproliferation och överlevnad. CREB (cAMP response element-binding protein) är en cellulär transkriptionsfaktor som kan öka gentranskriptionen mer än tio gånger. c-fos är ett protein med 380 aminosyror med en basisk leucinblixtlåsregion för dimerisering och DNA-bindning och en transaktiveringsdomän vid C-terminus. Det är involverat i viktiga cellulära händelser, inklusive cellproliferation, differentiering och överlevnad. ETS domain-containing protein (Elk-1) är ett protein som hos människor kodas av ELK1-genen. Elk-1 fungerar också som en transkriptionsaktivator.
– Onkogenes
ERK1/2-aktivering främjar metaplasi och tumörutveckling genom fosforylering av Bim och Bid och orsakar proteasomnedbrytning av Bim och sekretering av Bad till fosfoserinbindande proteiner och därmed inhiberar apoptos. I en separat väg fosforylerar ERK1/2-aktivering FOXO3a vid Ser 294, Ser 344 och Ser 425 och underlättar FOXO3a-MDM2-interaktionen. Denna interaktion ökar nedbrytningen av FOXO3a genom en MDM2-beroende ubiquitin-proteasomväg, vilket leder till tumörutveckling.
– Utveckling och differentiering
Signalering från receptortyrosinkinaser som aktiverar Ras/Raf/MAP-kinasvägen reglerar utveckling och differentiering av många typer av celler. Huvudsakligen genom aktivering av signaltransducerare och aktivatorer av transkriptionsproteiner (STATs).
– Cellcykel
Cdc25 aktiverar cyklinberoende kinaser genom att ta bort fosfat från rester i Cdk:s aktiva plats. Den är känd för att kontrollera övergångarna från G1 till S-fas och G2 till M-fas. Erk1/2 kan aktivera Cdc25-proteinet och i sin tur främja cellcykelns utveckling.
4. Reglering av Erk-signalering
Regleringen av Erk-signalering är mycket viktig för att vi ska kunna upprätthålla cellernas normala funktion. Det finns flera regleringsstrategier för cellen för att reglera Erk-signalvägen, främst inkluderar reglering genom återkopplingsslingor, genom upp- och nedströmsställningar, genom fosfatas och hämmare av Erk-signalvägen.
– Reglering av återkopplingsslingor
Det finns flera punkter med negativ återkoppling i ERK-signalvägen (figur 1). MEK är ett mål och kan hämmas genom ERK-fosforylering av MEK1/2 vid Thr292 och Thr212. Fosforyleringen förhindrar ytterligare förstärkning av MEK-aktiviteten genom PAK1 och minskar därmed aktiveringen av ERK. Fosforylering av Raf på flera ställen av ERK utgör en andra möjlig återkopplingsmekanism. Hyperfosforylering av dessa platser förhindrar Rafs interaktion med Ras GTPas och främjar dess defosforylering av fosfataset PP2A. Aktivering eller transkriptionell uppreglering av fosfataser av ERK är en alternativ negativ återkopplingsmekanism. Dessa olika ERK-aktiverade negativa återkopplingsslingor utgör ett sätt att begränsa signalens varaktighet och återföra signalvägen till basaltillståndet.
– Upp- och nedströmsreglering av skelettproteiner
Skelettproteiner binder två eller flera komponenter i en signalväg för att föra dem i nära anslutning till varandra och på så sätt underlätta deras funktionella interaktion. Dessutom kan scaffolds rikta dessa signaleringsmoduler med flera enzymer mot olika cellplaceringar och därigenom öka fosforyleringen av en specifik undergrupp av substrat i nedströmsledet. Scaffolds påverkar också signalens varaktighet, förhindrar överlappning mellan liknande vägar och kan koppla aktiveringen av signalmodulen till en specifik aktiverande signal uppströms.
– Fosfataser och hämmare
Aktiviteten hos MAPK-vägarna bestäms av de konkurrerande effekterna av kinaser uppströms och inhiberande fosfataser. MAPK-vägarna avslutas genom avfosforylering av deras komponenter av flera fosfataser. Flera serin/treoninfosfataser, t.ex. PP2A och PP2Ca, och tyrosinfosfataser, t.ex. PTP-SL och HePTP, har visat sig inaktivera MAPK:er.
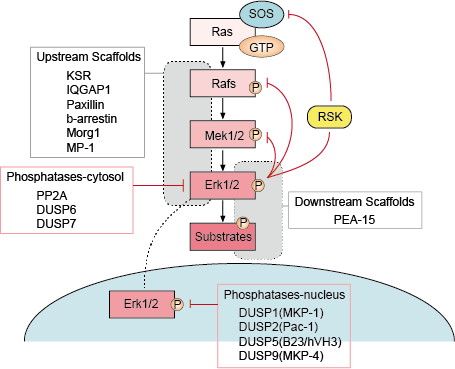
Figur 1. Reglering av Erk-signalvägen
5. Samband med sjukdomar
Sammanhängande med deras kritiska roller i viktiga cellaktiviteter, inklusive cellproliferation, differentiering och överlevnad eller död, har MAPK-signalvägarna involverats i patogenesen för många mänskliga sjukdomar.
– Alzheimers sjukdom
Alzheimers sjukdom (AD) är en neurodegenerativ sjukdom som kännetecknas av kognitiva störningar och minnesstörningar som tros bero på att det i hjärnan bildas både senila plack som innehåller amyloid-β (Aβ) och neurofibrillära trassel som innehåller det mikrotubuli-associerade proteinet tau. Bevis tyder således på att MAPK-signalvägar kan bidra till AD:s patogenes genom reglering av neuronal apoptos, β- och γ-sekretasaktivitet och fosforylering av APP och tau.
– Parkinsons sjukdom
Parkinsons sjukdom (PD) är den näst vanligaste neurodegenerativa sjukdomen. Olika observationer tyder på att MAPK-signalvägar bidrar till neuroinflammatoriska reaktioner och neurondöd som utlöses av α-synukleinaggregat eller funktionella brister i parkin eller DJ-1 i patogenesen för Parkinsons sjukdom.
– MAPK-signalering i cancer
Många av de cancerassocierade mutationerna av komponenterna i MAPK-signalvägarna har hittats i Ras och B-Raf, som båda deltar i ERK-signalvägen. ERK-signalvägen spelar en roll i flera steg i tumörutvecklingen. ERK:s fosforylering av proteiner som myosin light chain kinase, calpain, focal adhesion kinase och paxillin främjar cancercellernas migration. Vidare reglerar ERK1/2-signalering aktiviteten och nivåerna av proteiner ur Bcl-2-familjen, t.ex. det proapoptotiska proteinet BIM och det antiapoptotiska proteinet MCL-1, vilket främjar cancercellernas överlevnad.
| Keshet Y, Seger R. The MAP kinase signaling cascades: a system of hundreds of components regulates a diverse array of physiological functions. MAP Kinase Signaling Protocols: Second Edition, 2010: 3-38. | |
| Wortzel I, Seger R. The ERK cascade distinct functions within various subcellular organelles. Genes & cancer, 2011, 2(3): 195-209. | |
| Ramos J W. The regulation of extracellular signal-regulated kinase (ERK) in mammalian cells. The international journal of biochemistry & cell biology, 2008, 40(12): 2707-2719. | |
| Shaul Y D, Seger R. The MEK/ERK cascade: from signaling specificity to diverse functions. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2007, 1773(8): 1213-1226. | |
| Yao Z, Seger R. The ERK signaling cascade—views from different subcellular compartments. Biofactors, 2009, 35(5): 407-416. |